DIVERSIDAD Y ESTRUCTURA DE BOSQUES CONTRASTANTES EN LA REGIÓN DEL CHOCÓ-DARIÉN, COLOMBIA
DIVERSITY AND STRUCTURE OF CONTRASTING FORESTS IN THE CHOCÓ-DARIÉN REGION OF COLOMBIA
Luisa Fernanda Casas Herrera1*
Diana María González Flórez2
Beatriz Segura3
Ramón Antonio Mosquera4
Esteban Álvarez Dávila5
1 MSc, ECAPMA, UNAD Nodo Zona Occidente CEAD Medellín-Colombia
2Estudiante Ingeniería Agroforestal, ECAPMA, UNAD Nodo Zona Occidente CEAD Medellín-Colombia.
3 Estudiante Ingeniería Agroforestal, ECAPMA, UNAD Nodo Zona Occidente CEAD Turbo-Colombia.
4 PhD, ECAPMA, UNAD Nodo Zona Occidente CEAD Turbo-Colombia.
5PhD, ECAPMA, UNAD Nodo Zona Centro CEAD Bogotá-Colombia.
* luisa.casas@unad.edu.co
Resumen/ Abstract/ Resumo
El Chocó biogeográfico (ChB) es considerado un Hotspot de biodiversidad, pero existen pocos estudios comparativos de las diferencias regionales asociadas con la fisiografía o los suelos. En este estudio contrastamos la diversidad, abundancia y dominancia de especies entre bosques de “tierra firme vs pantanosos” en dos sitios del ChB. El primero ubicado en la costa pacífica (Jardín Botánico del Pacífico) y el segundo en la costa atlántica en el Darién (reserva Sasardí). Nuestros resultados concuerdan con otros estudios que muestran grandes diferencias entre bosques cercanos de tierra firme y pantanosa. Encontramos que dentro de cada región, las zonas inundables mostraron mayor número de individuos, pero menores valores de área basal, que las zonas de tierra firme. Esto se debe básicamente a que las zonas pantanosas están dominadas por especies de palmas, en particular por Euterpe oleracea Mart., con muchos individuos pero que alcanzan pequeños diámetros del tronco, generalmente menores de 15 cm. En relación con la diversidad alfa, los valores de riqueza de especies y diversidad de Shannon de bosques pantanosos presentan valores más bajos que los bosques de tierra firme en ambos sitios. El índice de dominancia de Simpson muestra un patrón contrario, y las curvas de rarefacción se corresponden con este patrón. La diversidad beta es alta, separando claramente los bosques de Tierra firme de los pantanosos dentro de cada región, y los de Tierra Firme entre las regiones. Se destaca la importancia de los bosques pantanosos, que si bien tienen una extensión reducida en el norte del Chocó, albergan un gran número de individuos de especies útiles para las comunidades. Además, prestan otros servicios ecosistémicos de importancia como el control de inundaciones y el almacenamiento de carbono.
Palabras clave: Chocó biogeográfico, bosques inundados, hiperdominancia, servicios ecosistémicos.
ABSTRACT
Biogeographic Choco (ChB) is a biodiversity hotspot, but there are few comparative studies of the regional differences associated with physiography or soils. In this research, we compared the diversity, abundance and dominance of species among "mainland vs. swamp" forests at two ChB sites. One of the sites is located in the Darien-Caribbean region (Sasardi reserve) and the other on the Pacific coast. The results confirm those of other studies that show large differences between nearshore and swamp forests. We found that within each region, the swamp areas showed higher numbers of individuals, but lower basal area values, than the mainland areas. The swampy areas are dominated by palm species, in particular Euterpe oleracea Mart., with many individuals but with small trunk diameters, generally less than 15 cm. In relation to alpha diversity, the species richness and Shannon diversity values of the swamp forests are lower than those of the mainland forests at both sites. Simpson's dominance index shows an opposite pattern, and the rarefaction curves correspond to this pattern. Beta diversity is high, clearly separating the forests of the mainland from the swamps within each region, and those of the mainland between the regions. The importance of the marshy forests, which, although small in size in the north of Choco, are home to a large number of individuals of species useful to the communities, is highlighted. In addition, they provide other important ecosystem services such as flood control and carbon storage.
Key words: Biogeographical Chocó, flooded forests, hyperdominance, ecosystem services.
Resumen Grafico
Introducción
El Chocó Biogeográfico que recibe más de 7000 mm de lluvia por año, es una región del neotrópico ubicada al noroccidente de nuestro país; y alberga bosques muy húmedos y pluviales tropicales (Rangel, 2004; Quinto & Moreno, 2014; Langendoen & Gentry, 1991), donde se presentan especies del centro y del sur del continente americano (Galeano et al. 1998; Ramirez 2011). El litoral pacífico de Colombia representa a nivel mundial un hotspot de conservación debido a su alta biodiversidad y endemismo (Myers et al. 2000). También hace un aporte significativo a la economía colombiana, ya que proporciona el 40% de la madera que se consume anualmente en el país. No obstante, a pesar de su importancia, estos recursos están amenazados por la deforestación y la degradación de los bosques (Melo & Cruz, 2003; IDEAM, 2017).
De acuerdo con varios estudios realizados en esta región (Langendoen & Gentry, 1991; Galeano, 2002; Torres et al. 2016; Mosquera et al. 2007; Quinto y Mosquera, 2014), se ha encontrado que los bosques del Chocó tienen características particulares en estructura y composición florística. En particular, se presenta una alta densidad de árboles y un número comparativamente elevado de especies; así se han registrado en promedio entre 150 – 250 especies árboles por hectárea, lo cual ha permitido situar a esta región en el segundo lugar en cuanto a riqueza florística en Colombia después de la Amazonía (Rangel 2004).
En relación con estas características en biodiversidad que presenta el Chocó Biogeográfico y la amenaza que representa la extracción desmedida del recurso maderero para la conservación de la misma, es necesario realizar estudios de su composición florística que induzcan a conocer la estructura y función de diferentes niveles jerárquicos; como también la exploración de posibles usos de las especies y los patrones de la distribución de la biodiversidad en el espacio y tiempo. Ésta información es primordial, para la gestión sostenible del territorio y para la toma de decisiones sustentadas científicamente; y es una necesidad urgente, que los investigadores, las instituciones y las naciones deben enfatizar para diseñar acciones de conservación (Villareal et al. 2006).
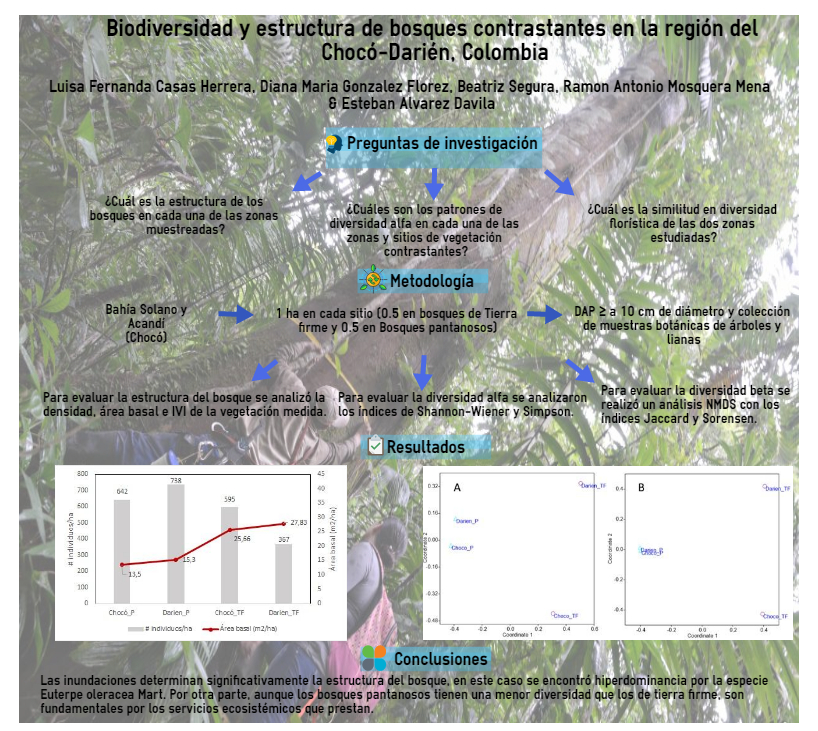
Con la presente investigación se espera contribuir al conocimiento de la composición florística en zonas con bosques primarios ubicados al norte del Chocó biogeográfico, en los municipios de Bahía Solano y Acandí. Para evaluar la diversidad, se recolectó la información en dos tipos de vegetación contrastante (bosque de tierra firme vs bosque pantanoso). Los objetivos principales buscan dar respuesta a las siguientes preguntas de investigación: ¿Cuál es la estructura de los bosques en cada una de las áreas muestreadas?¿Cuáles son los patrones de diversidad alfa en cada una de las zonas? y ¿Cuál es la similitud en diversidad de las dos zonas?
Materiales y métodos
Área de estudio
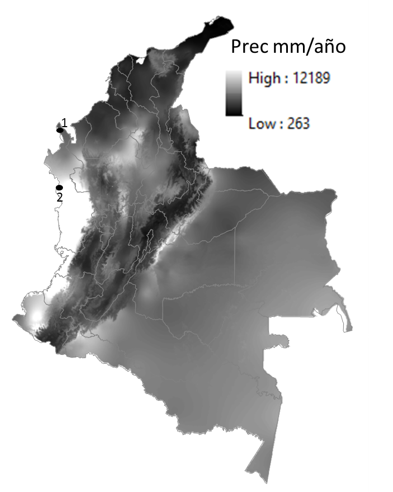
El estudio se llevó a cabo en bosques primarios de los municipios de Bahía Solano (Sitio 1 en el Jardín Botánico del Pacífico) y Acandí (Sitio 2 en la Reserva Sasardí) al norte del departamento del Chocó (Figura 1).
Figura 1. Localización en Colombia de los sitios de estudio. 1) Reserva Sasardí en el municipio de Acandí. 2) Jardín Botánico del Pacífico, en el municipio de Bahía Solano.
Fuente: www.worldclim.org
Sitio 1. Jardín Botánico del Pacífico
El Jardín Botánico del Pacífico está localizado en el municipio de Bahía Solano, en el departamento del Chocó, en la vereda Mecana en las coordenadas 06° 13’ 27” latitud norte y 77° 24’ 14” longitud oeste. Tiene una extensión total de 170 hectáreas de las cuales 120 corresponden a bosque. El jardín botánico está ubicado en la zona de vida bosque pluvial tropical (bp-T) según Holdrigde (2000). Su clima está definido por una temperatura media 26°c, una precipitación superior a 6.000 mm/año, un brillo solar entre 52 y 100 horas mensuales, una humedad relativa del 85% y una nubosidad entre 5 y 7 octas (IDEAM, MAVDT Y PNUD,2006; Torres, Mena & Alvarez, 2017).
Sitio 2. Reserva Sasardí
La reserva integral y Ecoaldea Sasardí (RIES) se encuentra ubicada en el municipio de Acandi en el departamento del Chocó, en la vereda Sasardí sobre la serranía costera de Tripogadí al occidente del golfo de Urabá-Chocó, tiene una extensión de 60 hectáreas donde cerca de la mitad de estas corresponde a bosque primario, localizada en las coordenadas 08° 22′ N y 77° 08′ O, a 129 msnm cuenta con una precipitación media de 2800 mm/año y 28 a 35 ºC de temperatura, ubicada en la zona de vida según Holdridge de bosque húmedo tropical bh-T.
Unidades experimentales y medición de variables
En cada uno de los sitios (Chocó o JBP y Darién o Sasardí) se realizó un inventario de 1 ha, incluyendo dos tipos de vegetación. 1) Bosques de tierra firme y 2) Bosques pantanosos sobre suelos permanentemente inundados. En cada tipo de vegetación/sitio se contó con un inventario de 0.5 ha. Las parcelas fueron establecidas durante el año 2014. De aquí en adelante se utilizarán códigos que describen los bosques de tierra firme y pantanosos en el JBP como Chocó_TF y Chocó_P, los mismos bosques en Sasardi como Darién_TF y Darién_P.
En ambas parcelas se rotularon y midieron todos los individuos de árboles y lianas ≥ a 10 cm de diámetro. Se midió el diámetro a 1.3 m del suelo. Para la identificación de las especies se hicieron colecciones botánicas, aunque algunas de fácil reconocimiento (por ej. Euterpe oleracea) fueron identificadas en campo. El trabajo de herborización y de identificación de las colecciones fue realizado en los herbarios del Jardín Botánico de Medellín (JBMED) y de la Universidad de Tolima (UT) que cuenta con un buen número de colecciones históricas de la región del Choco-Darién.
Análisis estadístico
Para evaluar la estructura del bosque se halló la abundancia, área basal e índice de valor de importancia (IVI) para cada uno de los sitios muestreados y tipos de vegetación. Para evaluar la diversidad alfa y beta, se utilizaron índices que ayudaron a resumir la información en un solo valor y que permitieron unificar cantidades para hacer comparaciones. Estos índices se calcularon por medio del programa estadístico R, en el cual se utilizó la librería Vegan.
Diversidad alfa (α): se utilizaron los siguientes estimadores basados en la abundancia relativa de especies. Estos índices combinan la riqueza y la abundancia relativa de especies.
Shannon-Wiener (H´): es un índice de equidad que tiene en cuenta la abundancia de cada especie y que tan uniformemente se encuentran distribuidas. Mide la heterogeneidad de la comunidad, el valor máximo será indicador de una situación en la cual todas las especies son igualmente abundantes (Villareal et al. 2006; Melo & Cruz, 2003; Moreno, 2001).
H’=
Donde:
H = Diversidad de Shannon
pi= (ni/N) Abundancia proporcional (relativa)
Índice de Simpson (D): es un índice de dominancia que se enfatiza en las especies más comunes o aquellas que están mejor representadas (dominan) en el área de muestreo. Éste índice se refiere a la probabilidad, de que dos individuos tomados al azar de una muestra, correspondan a la misma especie (Villareal et al. 2006; Melo & Cruz, 2003; Moreno, 2001).
D = ∑ pi2 o D = ∑ [ni (ni-1)/N(N-1)]
Donde:
pi = Abundancia proporcional
ni = Número de individuos de la iésima especie
N = Número de individuos totales
Diversidad Beta (β): se evaluó la diversidad β mediante los índices de Jaccard y Sorensen, para analizar las diferencias entre los hábitats y los cambios en la composición de las especies entre los sitios. Mientras menos especies compartan las dos áreas de estudio, mayor será su beta diversidad (Melo & Cruz, 2003).
Jaccard (IJ): relaciona el número de especies compartidas con el número total de especies exclusivas. El intervalo de valores para este índice va de 0 cuando no hay especies compartidas entre ambos sitios, hasta 1 cuando los dos sitios tienen la misma composición de especies (Villareal et al. 2006, Moreno, 2001).
IJ=
Donde:
a = número de especies presentes en el sitio A
b = número de especies presentes en el sitio B
c = número de especies presentes en ambos sitios A y B
Sorensen (IS): relaciona el número de especies compartidas con la media aritmética de las especies de ambos sitios. El intervalo de valores para este índice va de 0 cuando no hay especies compartidas entre ambos sitios, hasta 1 cuando los dos sitios tienen la misma composición de especies (Villareal et al. 2006, Moreno, 2001).
IS =
Donde:
a = número de especies presentes en el sitio A
b = número de especies presentes en el sitio B
c = número de especies presentes en ambos sitios A y B
Resultados y Discusión
Diversidad alfa, abundancia y área basal
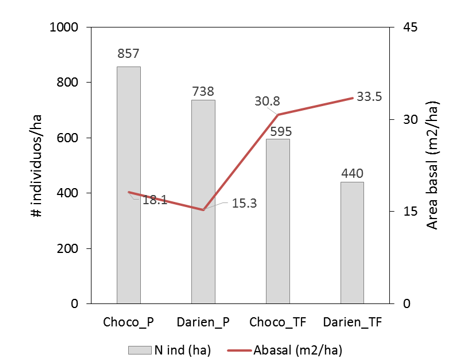
Se encontró un total de 1129 individuos en las 2 ha, de los cuales 575 son árboles, 547 son árboles palmas y 7 son lianas. Para los dos sitios de muestreo, los bosques pantanosos tienen una mayor densidad de árboles pero menor área basal que los bosques de tierra firme (Figura 2). Los 1129 individuo estuvieron representados por 130 especies, 100 géneros y 40 familias (ver listado en Anexo 1). La mayor riqueza y diversidad de especies se encontró en los bosques de tierra firme en comparación con los pantanosos, y los bosques de tierra firme del Darién tuvieron mayor riqueza y diversidad que los bosques del Chocó; un patrón contrario se encontró para el índice de dominancia de Simpson (Tabla 1).
Figura 2. Comparación del número de individuos y el área basal/ha en bosques de tierra firme (TF) y pantanosos en Choco y Darién.
Tabla 1. Comparación de la diversidad alfa. Riqueza de especies y la diversidad entre el Choco y el Darien, considerando bosques pantanosos (P) y bosques de tierra firme (TF).
| Indice | Chocó_P |
Darién_P |
Choco_TF |
Darien_TF |
Número especies |
9 |
17 |
59 |
63 |
Shannon_H |
0.38 |
1.66 |
3.44 |
3.62 |
Dominance_D (Simpson) |
0.87 |
0.30 |
0.05 |
0.04 |
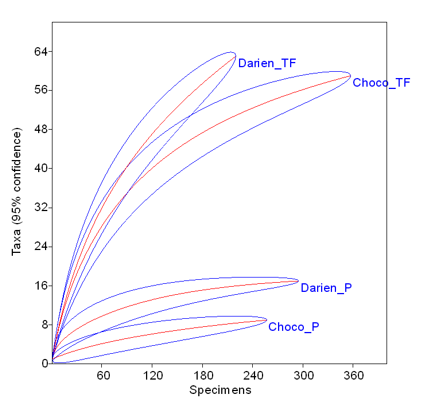
Los bosques de pantano están claramente dominados por las palmas, principalmente en el Chocó, y a nivel de las especies tienen una composición más homogénea que los bosques de tierra firme, con una baja riqueza y diversidad, y la presencia de una especie híper dominante, Euterpe oleraceae (Tabla 2). Con respecto a los bosques de tierra firme se encontraron especies con mayor dominancia en el Chocó, posiblemente como respuesta a perturbaciones pasadas, dada la alta dominancia de Jacaranda y Cecropia. Las 5 especies con mayor IVI representan el 89,7%, 80,1%, 41,6% y el 34.0% en Chocó_P, Darién_P, Chocó_TF y Darién_TF, respectivamente (Tabla 2). Las curvas de rarefacción muestran claramente que los bosques de tierra firme son significativamente más ricos en especies que los pantanosos, y los del Darién más ricos que los del Chocó (Figura 3).
Tabla 2. Especies dominantes en los diferentes tipos de bosque.
N |
ESPECIE |
Abun ab |
Abas ab |
Frec ab |
Abun rel |
Abas rel |
Frec rel |
IVI % |
Bosque Pantanoso del Chocó |
||||||||
1 |
Euterpe oleracea |
239 |
3.07 |
8 |
93.0 |
56.4 |
42.1 |
63.8 |
2 |
Jacaranda copaia |
7 |
0.95 |
2 |
2.7 |
17.4 |
10.5 |
10.2 |
3 |
Pourouma sp |
2 |
0.35 |
2 |
0.8 |
6.5 |
10.5 |
5.9 |
4 |
Virola reidii |
2 |
0.57 |
1 |
0.8 |
10.5 |
5.3 |
5.5 |
5 |
Exarata chocoensis |
3 |
0.04 |
2 |
1.2 |
0.8 |
10.5 |
4.2 |
Bosque Pantanoso del Darien |
||||||||
1 |
Euterpe oleracea |
136 |
1.42 |
10 |
46.1 |
23.3 |
14.5 |
28.0 |
2 |
Manicaria saccifera |
79 |
2.17 |
10 |
26.8 |
35.5 |
14.5 |
25.6 |
3 |
Raphia taedigera |
23 |
1.17 |
9 |
7.8 |
19.2 |
13.0 |
13.3 |
4 |
Mauritiella macroclada |
20 |
0.28 |
10 |
6.8 |
4.5 |
14.5 |
8.6 |
5 |
Sterculia pilosa |
5 |
0.31 |
5 |
1.7 |
5.0 |
7.2 |
4.7 |
Bosque de Tierra firme del Choco |
||||||||
1 |
Jacaranda copaia |
43 |
4.42 |
14 |
12.0 |
28.7 |
6.4 |
15.7 |
2 |
Cordia sp |
47 |
1.07 |
14 |
13.2 |
7.0 |
6.4 |
8.8 |
3 |
Cecropia schreberiana |
22 |
1.28 |
9 |
6.2 |
8.3 |
4.1 |
6.2 |
4 |
Brosimum alicastrum |
16 |
1.23 |
10 |
4.5 |
8.0 |
4.5 |
5.7 |
5 |
Chrysochlamys sp |
25 |
0.56 |
11 |
7.0 |
3.7 |
5.0 |
5.2 |
Bosque de Tierra firme del Darien |
||||||||
1 |
Pouteria amygdalicarpa |
26 |
1.39 |
13 |
11.8 |
8.3 |
7.9 |
9.3 |
2 |
Cavanillesia platanifolia |
2 |
3.26 |
2 |
0.9 |
19.5 |
1.2 |
7.2 |
3 |
Batocarpus costaricensis |
19 |
1.01 |
11 |
8.6 |
6.0 |
6.7 |
7.1 |
4 |
Anacardium excelsum |
2 |
2.27 |
2 |
0.9 |
13.5 |
1.2 |
5.2 |
5 |
Castilla tunu |
9 |
1.39 |
5 |
4.1 |
8.3 |
3.0 |
5.1 |
Figura 3. Curvas de rarefacción (número de individuos vs número de especies) para los bosques estudiados (líneas rojas) con sus correspondientes intervalos de confianza (líneas azules). Se muestran las curvas para bosques pantanosos y bosques de tierra firme.
Diversidad beta
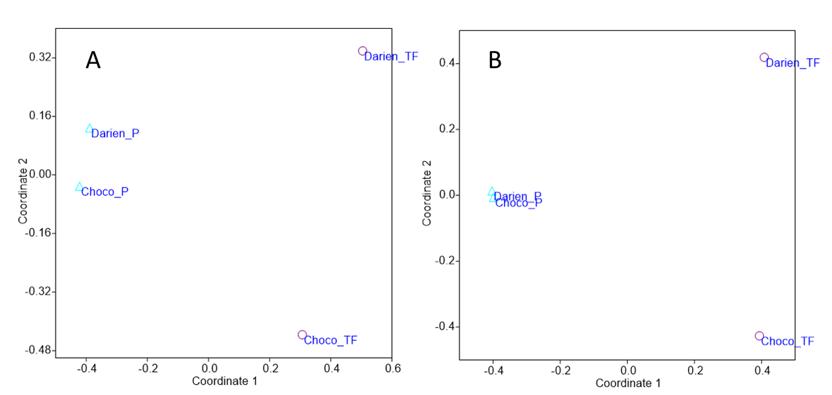
No se encontró ninguna especie en común entre los diferentes tipos de bosque. Los bosques de tierra firme de Chocó y Darién comparten solo el 6.4% de las especies mientras que los pantanosos comparten el 13% entre las dos regiones (Tabla 3). El análisis NMDS (ordenamiento indirecto) muestra claramente una separación florística entre los pantanosos y los bosques de tierra firme. Adicionalmente, el análisis NMDS indica que existen grandes diferencias florísticas entre los bosques de tierra firme en Chocó y Darién, y una alta similitud entre los bosques pantanosos de las dos regiones (Figura 4).
Tabla 3. Número y porcentaje de especies comunes entre los diferentes tipos de bosque.
Chocó_TF |
Darién_P |
Chocó_P |
|
# especies comunes |
|||
Darién_TF |
7 |
1 |
0 |
Chocó_TF |
1 |
2 |
|
Darién_P |
3 |
||
% especies comunes |
|||
Darién_TF |
6.4 |
1.3 |
0.0 |
Chocó_TF |
1.4 |
3.2 |
|
Darién_P |
|
|
13.0 |
Figura 4. Diversidad Beta. Ordenamiento multidimensional métrico (NMDS) de los bosques pantanosos y de tierra firme en el Choco y el Darién, utilizando diferentes índices de similitud: (A) Jaccard y (B) Sorensen.
DISCUSIÓN
La mayor parte de los estudios en regiones tropicales que muestran las diferencias entre bosques de tierra firme y pantanosos, se han realizado en la cuenca del rio Amazonia (Anexo 3; ver además revisiones recientes en Pitman et al. 2014 y Wittmann et al. 2017). Como en nuestro caso, la hiperdominancia en los pantanos arbolados ha sido reportada tanto en bosques tropicales como templados. Henderson et al. (1997) reportan que Euterpe oleracea Mart. es una palma con origen en los trópicos y sub trópicos de Suramérica, ocupando generalmente terrenos inundados y esporádicamente suelos de tierra firme (Cavalcante, 1991). La E. oleracea crece a lo largo de la costa del Pacífico desde Panamá (San Blas) hasta el norte de Ecuador (Esmeraldas). También se encuentra en áreas estuarinas de Trinidad, Venezuela (delta del Orinoco), las Guayanas y Brasil (Estado Amazónico). En Colombia, crece también en el valle medio e inferior del río Atrato, en el bajo Cauca y en el valle medio del río Magdalena, y en el alto Sinú (Galeano y Bernal, 2010). Estudios desarrollados en el estuario amazónico, como el de Cattanio et al. (2004), plantean que E. oleracea es una de las especies mejor adaptadas a suelos inundables gracias a sus neumatóforos.
Dada la importancia de estos bosques, existen desde hace décadas trabajos enfocados en entender su distribución espacial a nivel continental (Klinge et al. 1990) y su valor para la seguridad alimentaria de las comunidades (Kahn, 199; Montenegro-Gómez, S. P., & Rosales-Escarria, M. 2015). Más recientemente, la importancia de los bosques pantanosos viene siendo reconocida por otros servicios ecosistemicos; por ejemplo, se sabe actualmente que los bosques tropicales en general son importantes reservas de carbono, pero la contribución de los bosques pantanosos de agua dulce a esta reserva no se ha cuantificado en gran medida (Igu y Marchant, 2016). Más aún, recientemente se han reportado grandes extensiones de suelos pantanosos en amazonia que almacena grandes cantidades de carbono (Draper et al. 2014).
En cuanto a la riqueza florística encontrada en los bosques del Chocó y bosques del Darién; sería pertinente evaluar en eventuales investigaciones los factores ecológicos que se podrían correlacionar con la diversidad presente en cada una de los sitios; así por ejemplo, Gentry (1982), llegó a la conclusión en un estudio realizado sobre diversidad de especies en bosques neotropicales, que la diversidad aumentaba con la cantidad y la estabilidad de la precipitación y, en menor grado, con la fertilidad del suelo.
Por otra parte, también sería pertinente evaluar la riqueza florística de los bosques, teniendo en cuenta los árboles, palmas y lianas con DAP menores a 10 cm, pues aunque los estudios centrados en el dosel del bosque, determinan en gran parte su estructura y funcionamiento (Berry, 2002); en algunos estudios en bosques del Chocó se ha encontrado que la riqueza de especies está representada principalmente por el componente herbáceo y en especial por las especies de plantas epífitas (Galeano et al. 1998).
Con respecto a las diferencias en la composición florística que se encontraron entre los bosques evaluados, se observa las mismas características que se han encontrado cuando se evalúan los patrones de diversidad beta entre bosques neotropicales de bajura; por lo general, los bosques inundables son menos diversos que los bosques de tierra firme y ambos tienen relativamente pocas especies en común (Berry, 2002).
De acuerdo con lo anterior, en este estudio se pudo observar que los dos índices evaluados (Jaccard (Ij) y Sorensen (Is)) estiman el recambio florístico entre sitios de muestreo de manera similar, donde se encuentra que generalmente entre todos los sitios la composición florística es muy diferente, ya que los valores obtenidos son muy cercanos al número cero. No obstante y de acuerdo con las leves diferencias en los valores obtenidos, se analiza que entre los bosques pantanosos de los dos sitios de muestreo, existe una mayor similitud en la composición florística en comparación con los bosques de tierra firme de los dos sitios. Además, entre los dos sitios de muestreo y tipos de bosques diferentes (pantanosos y de tierra firme) existe alta heterogeneidad en la riqueza de especies presentes, ya que se obtuvieron los menores valores para los índices evaluados.
Conclusiones
La conclusión más obvia de estos dos muestreos, y otros hechos en los mismos tipos de bosque, es que las inundaciones determinan significativamente la estructura del bosque. Esto se puede observar en la densidad total del tallo, el área basal y la diversidad. Si bien los bosques pantanosos tienen una menor diversidad que los de tierra firme son fundamentales por los servicios ecosistémicos que prestan. No solo contribuyen a controlar las inundaciones, sino que también tienen una alta densidad de individuos útiles para las comunidades y almacena grandes cantidad de carbono, especialmente en el suelo.
Literatura citada