PRESENCIA DE ESPORAS DE HONGOS MICORRÍZICO ARBUSCULARES EN CINCO ESTADOS SUCESIONALES DE BOSQUE ALTOANDINO COLOMBIANO
PRESENCE OF ARBUSCULAR MYCORRHIZAL FUNGI SPORES IN FIVE SUCCESSION STATES OF ALTOANDINO COLOMBIAN FOREST
Favio López Botía1
Diana Yulieth Peña2
1 Biólogo. MSc. Docente Proyecto Curricular de Ingeniería Forestal. Facultad del Medio Ambiente y Recursos Naturales. Universidad Distrital Francisco José de Caldas. Bogotá Colombia.
falopezb9@hotmail.com
2Estudiante Ingeniera Forestal. Universidad Distrital Francisco José de Caldas. Bogotá Colombia.
dianay.ps@gmail.com
Resumen
Dadas las condiciones de ubicación geográfica de Colombia en la franja tropical, el país cuenta con una alta diversidad de ecosistemas. El bosque natural representa un 52% de la superficie total del territorio nacional y se ha comprobado la alta riqueza que posee en lo referido a especies vegetales, animales y de hongos; sin embargo, la información acerca de la biología del suelo en el bosque natural es incipiente y gran cantidad de especies no se han clasificado taxonómicamente, llevando al desconocimiento de microorganismos como los hongos micorrízico arbusculares (HMA), importantes en el establecimiento de comunidades vegetales y ecosistemas. Se realizó el aislamiento de esporas de HMA en muestras compuestas de suelo tomadas en la rizósfera de cinco estados sucesionales de bosque altoandino; posteriormente se hizo el reconocimiento y conteo de esporas y fue evaluado el efecto de las condiciones físicoquímicas del suelo de cada estado sucesional en la densidad de esporas. El análisis estadístico consistió en un análisis de componentes principales y la representación gráfica de un análisis de agrupamiento. Se registró la presencia de esporas de HMA en todas las etapas sucesionales del bosque altoandino, siendo el estado maduro el que mayor densidad tuvo con un promedio de 20 esporas*g-1; la compactación resultó ser una de las características con mayor influencia en la variación de la densidad de esporas y se observó que aspectos como la concentración de metabolitos secundarios en las raíces de las especies vegetales pueden influenciar la esporulación y por ende la densidad de esporas de los HMA.
Palabras clave: Bosque Andino, Compactación, Espora, Materia orgánica, Simbiosis.
Abstract
Given the geographic location conditions of Colombia in the tropical belt, the country has a high diversity of ecosystems. The natural forest represents 52% of the total surface of the national territory and it has been proven that it has a high level of wealth in terms of plant, animal and fungal species; However, information about the biology of the soil in the natural forest is incipient and a large number of species have not been classified taxonomically, leading to the lack of knowledge of microorganisms such as arbuscular mycorrhizal fungi (AMF), important in the establishment of plant communities and complex ecosystems. The isolation of HMA spores was do in composite samples of soil taken from the rhizosphere of five successive high-andean forest states; Subsequently, spore recognition and counting was carried out and the effect of the physicalchemical soil conditions of each successional state on the spore density was evaluated. The statistical analysis consisted of an analysis of the main components and the graphic representation of a grouping analysis. The presence of AMF spores was recorded in all the successional stages of the high Andean forest, with the mature state having the highest density with an average of 20 spores * g-1; compaction turned out to be one of the characteristics with the greatest influence on the variation of spore density, and it was observed that aspects such as the concentration of secondary metabolites in the roots of plant species can influence sporulation and therefore the spore density of the AMF.
Keywords: Andean Forest, Biodiversity, Compaction, Organic matter, Spore, Symbiosis.
Introducción
Colombia es uno de los países con mayor diversidad biológica del planeta, se ubica en el primer lugar de diversidad en aves y orquídeas, segundo en número de especies de plantas, peces dulce-acuícolas y anfibios; tercero en diversidad de reptiles y palmas, y cuarto en diversidad de mamíferos (SIB, 2014). Del total de la biodiversidad mundial, aproximadamente el 75% de ella se concentra en solo diecisiete países, siendo Colombia uno de los más representativos (Lesmes, 2013; WWF, 2017). La gran variedad de ecosistemas forestales con los que cuenta, sustentan la megadiversidad de la nación, la cual constituye el 10 % de la biodiversidad mundial (Colciencias 1999).
Las tierras cubiertas por bosque natural ocupan el 52% de la superficie continental nacional, que corresponde a aproximadamente 60 millones de hectáreas, ubicándolo como el tercer país de Suramérica con mayor área de bosques naturales; dichos bosques se distribuyen a lo largo de las cinco regiones biogeográficas del país, pero el mayor porcentaje se concentra en las regiones Amazónica y Andina, que en conjunto representan el 84,7% de dicha área (MINAMBIENTE, 2017). Debido a la alta diversidad del país, una gran cantidad de especies no se han descrito ni clasificado taxonómicamente, siendo la biología del suelo una de las áreas menos estudiada.
El bosque altoandino está caracterizado por la presencia de varios estratos: comenzando por un manto de plántulas de especies confinadas a la parte inferior del bosque, seguido de plantas poco lignificadas pertenecientes al sotobosque, herbáceas, reptantes y/o de bajo porte hasta una bóveda o dosel formado por árboles de altura considerable, en cuyas copas frondosas se albergan otras especies animales y vegetales (IDEAM, 1997).
En Colombia, los procesos de cambio de uso de la tierra han llevado a la continua deforestación generando daños que resultan ser frecuentemente irreversibles (DANE, 2015). Para el caso del bosque altoandino, este es considerado uno de los ecosistemas más amenazados a causa de la expansión de la frontera agrícola, ya que ofrece condiciones favorables para el asentamiento humano, tales como: clima, aptitud del suelo, flora y fauna; consecuencia de ello es que en esta región, que representa el 27% de la extensión territorial del país, se encuentre asentada el 41% de la población colombiana (Andrade, 1993).
El impacto constante de labores “más productivas”, ha originado la transformación de los ecosistemas, la disminución de la biodiversidad, entre otras consecuencias que llevan a que del bosque altoandino actualmente existan relictos de vegetación, siendo uno de los ecosistemas donde se registran las mayores tasas de pérdida de diversidad biológica (Avella et al., 2014; Cantillo et al., 2005). Sumado a lo anterior, factores como la erosión del suelo que afecta al 35 % del territorio nacional con más de 4.300.000 ha clasificadas con erosión severa y muy severa (IGAC, 2002), incrementan la vulnerabilidad del bosque altoandino y de la biodiversidad que sustenta. Debido a ello, es necesario desarrollar medidas que puedan hacer frente a las alteraciones que siguen presionando a este ecosistema.
En los programas de restauración de ecosistemas perturbados, además de los componentes vegetales por encima del suelo, se evalúan las condiciones biológicas y microbianas del mismo (Hernández & Pastor, 2008). Se han ejecutado prácticas de recuperación en las que se realiza la inoculación de hongos micorrízicos a plantas germinadas en vivero demostrando que dichas plantas tienen mayor capacidad de establecimiento y supervivencia, en comparación con aquellas que no presentan asociación con estos hongos (Cuenca et al., 1998; Falcon et al., 2016; Medina-Garcia, 2016).
Los hongos micorrízicos se destacan por cumplir diversas funciones dentro de la dinámica suelo-planta, una de sus funciones principales, es extender el área de absorción de nutrientes de las raíces en el suelo, haciendo más eficiente el proceso, especialmente importante en ambientes desfavorables (Azcón & Barea, 1992; Yang et al., 2014). Se ha demostrado que la superficie de absorción de las raíces colonizadas con micorrizas se incrementa hasta en 1.000 veces (Harley & Smith, 1983). Gracias a ello, las plantas micorrizadas presentan una mayor tolerancia ante la sequía, altas temperaturas, metales pesados, salinidad, toxinas (principalmente Cianotoxinas y micotoxinas) (Hart & Trevors, 2005) y a la acidez del suelo (Moncayo, 2002; Rangel et al., 2013; Cardona et al., 2017). Por otro lado, los hongos micorrízicos arbusculares permiten la agregación de las partículas del suelo mejorando su estructura mediante la secreción de glomalina por parte de hifas extrarradicales (Morell et al., 2009; Lehmann & Rillig, 2017), influencian la sucesión de comunidades de plantas (Janos, 1980) y su competitividad (Allen & Allen, 1986).
El término latino micorriza, cuyo significado es “raíz fungosa”, fue propuesto por el fitopatólogo alemán A.B. Frank en 1885 para describir globalmente toda una serie de estructuras que forman la gran diversidad de asociaciones que se establecen entre varios géneros de hongos del suelo que pertenecen al reino Fungi, y las raíces de la mayor parte de las plantas vasculares y algunas no vasculares (Gómez et al., 2007; Harley & Smith, 1983; Redecker et al., 2013). Se ha registrado la presencia de estos organismos desde hace más de 450 millones de años (Bonfante & Desiro, 2015) siendo el resultado de la coevolución mantenida entre las raíces de plantas y hongos micorrízicos como parte del avance colonizador de las plantas acuáticas primitivas hacia el medio ambiente terrestre (Honrubia, 2009). Actualmente se sabe que esta relación simbiótica se da en aproximadamente el 80% de la especies vegetales así como en el 92% de las familias de plantas (Blackwell, 2011). Es por ello que Moncayo (2002), expresa que la presencia de micorrizas en una planta es la norma y no la excepción.
Según Gómez et al., (2007) una de las razones por la cual la asociación simbiótica con micorriza ha despertado gran interés, es por su relación con la absorción del fósforo insoluble en el suelo por parte de las plantas colonizadas, ya que representa una de las alternativas viables para disminuir la deficiencia de este elemento que limita el desarrollo agropecuario y silvícola en suelos ácidos del trópico (Pedraza, 1981; Cabrales et al., 2016; Luna et al., 2016; Díaz, 2016).
La relación simbiótica planta-hongo supone un beneficio también para el hongo, que consiste en el suministro de compuestos orgánicos y de un hábitat protegido por parte del macrosimbionte; según Graham (2000) el hongo necesita alrededor de un 20% del producto fotosintético para su adecuado establecimiento, lo cual implica un costo para la planta, no obstante, este gasto se ve compensado por las ventajas que trae la asociación al macrosimbionte (Blaszkowski, 2007; Valdez & Vargas, 2011).
De acuerdo con lo anterior se constituye como objetivo de investigación evaluar la presencia de hongos micorrízicos arbusculares en suelos de cinco estados sucesionales del bosque altoandino en Guasca - Cundinamarca, así como determinar el efecto de las condiciones físicoquímicas edáficas en el establecimiento de dichos hongos, encaminado a dar las pautas para identificar los complejos ecológicos a nivel microbiano en los suelos del ecosistema analizado.
Materiales y métodos
Fase de campo
La fase de campo se realizó en jurisdicción del municipio de Guasca, departamento de Cundinamarca, dentro de la reserva forestal Carpatos, entre los 2600 y 3000 m de altitud, precipitación media de 1977 mm año-1, temperatura media de entre 10 y 12 ºC. Los suelos de la región son jóvenes con alta presencia de cenizas volcánicas y materia orgánica (Cantillo et al., 2005). Se tomaron muestras compuestas de suelo en cinco zonas dentro del área evaluada, cada una realizada en sitios representativos ecológicamente dentro del ecosistema, y en ecotonos que garantizaran captar la variación de la relación suelo – planta, entre los diferentes estados sucesionales establecidos en los cuales se identificaron cinco asociaciones vegetales, como se muestra en la tabla 1.
El área de muestreo corresponde a una extensión de 558 ha, dentro de las cuales 373 ha estan cubiertas por bosques representativos de la flora andina y 185 ha por pastos Kikuyo y Poa (Pennisetum clandestinum y Holcus lanatus) (Cantillo et al., 2005). En cada una de las asociaciones se establecieron tres puntos de muestreo, en las que se tomaron 3 submuestras al azar, de suelo entre 0 y 20 cm de profundidad en zona de influencia de raíces (rizósfera); en total se tomaron 9 submuestras por asociación que corresponden a 3 muestras compuestas y 45 submuestras para el total del área muestreada, que corresponden a 15 muestras compuestas. Para cada asociación se recolectaron datos tales como ubicación geográfica y altitud con GPS Garmin Etrex 10; temperatura del suelo con termómetro de tierra Greenco; temperatura ambiente con termohigrómetro digital y resistencia del suelo con penetrómetro Pocket penetrometer CL – 700A SOIL TEST INC. USA, con escala entre los 0.25 y 4.5 kg/cm2
Para la selección de los estados sucesionales se tuvo en cuenta que el área de estudio corresponde a un bosque secundario tardío, por lo cual se consideraron características fisionómicas y fitosociológicas de las comunidades vegetales allí presentes, a fin de tener una posición objetiva en la determinación de cada estado. El estado sucesional pionero se caracteriza por especies heliófitas de rápido crecimiento y una alta homogeneidad; el intermedio temprano presenta especies heliófitas y propágulos de hemiesciófitas manteniendo la alta homogeneidad del paisaje; en el estado intermedio existen especies hemiesciofitas de mayor porte y disminuye la abundancia de heliófitas; el estado intermedio tardío ya cuenta con un mosaico donde predominan especies hemiesciófitas e inicia el desarrollo de especies esciófitas, hay alta heterogeneidad. Finalmente, el estado maduro alcanza el máximo desarrollo sucesional con menor presencia de especies hemiesciófitas y mayor desarrollo de esciófitas. Cada uno de los sitios de muestreo fue debidamente georreferenciado, ubicándose el más bajo a 2820 msnm, mientras que el de mayor altura fue muestreado a los 2870 msnm.
Tabla 1. Estados sucesionales determinados en el bosque altoandino evaluado.
| ESTADO SUCESIONAL | ASOCIACION |
DESCRIPCION |
Pionero |
I |
Weinmannio pinnatae - Miconietum theaezantis |
Intermedio Temprano |
II |
Ocoteo callophyllae - Weinmannietum pinnatae |
Intermedio |
III |
Clusio multiflorae - Weinmannietum balbissianae |
Intermedio Tardío |
IV |
Rubo floribundi - Pteridietum aquilini |
Maduro |
V |
Hedyosmo crenati - Myrcinetum coriaceae |
Fuente de: Adaptación de Silva et al., 2006.
Fase de Laboratorio
El procesamiento y análisis de las muestras obtenidas en campo se realizó en los laboratorios de Fisiología Vegetal y Suelos, adscritos al proyecto curricular de Ingeniería Forestal, sede El Vivero de la Universidad Distrital Francisco José de Caldas. Se realizó el correspondiente análisis de características físicoquímicas del suelo proveniente de cada sucesión vegetal, evaluando: pH con potenciómetro (AOAC. 1990); compactación con penetrómetro PocketCL- 700A, materia orgánica según método de Walkley-Black (Kateman y Buydens, 1993), fósforo disponible con Bray II (Fassbender, 1975); fósforo en solución con resina de intercambio catiónico (AOAC, 1990); calcio, magnesio, potasio, hierro, zinc y boro siguiendo las metodologías señaladas en AOAC (1990).
El aislamiento de esporas se realizó para tres repeticiones de cada muestra compuesta, el procedimiento consistió en tomar 20 gramos de suelo fresco y realizar la extracción de esporas mediante la técnica de tamizado húmedo y decantado propuesto por Gerdemann & Nicolson (1963) así como centrifugado en gradiente de sacarosa de Walker et al. (1982) el cual corresponde a una adaptación del procedimiento de Gerdemann & Nicolson (1963), con variables prácticas realizadas por Gemma (1989). Una vez aisladas las esporas, se realizó el conteo total de aquellas que tenían características de turgencia y citoplasma intacto.
Análisis de datos
Para el análisis de los datos se utilizó el paquete estadístico R project © versión 3.5.1 mediante el cual se efectuó una evaluación descriptiva inicial y posteriormente ANAVA de una vía con cada una de las variables fisicoquímicas y la abundancia de esporas, luego se aplicó el test de Tukey (HSD) con nivel de significancia α=5%. A continuación se realizó un análisis de componentes principales y la representación gráfica de un análisis de agrupamiento empleando la librería FactoMineR.
Resultados y Discusión
En la tabla 2 se registran los datos obtenidos, del suelo de cada uno de los sitios muestreados, en cuanto a sus características fisicoquímicas, así como el número de esporas por gramo encontradas en un rango altitudinal de entre los 2820 a 2870 m de altitud.
Tabla 2. Características fisicoquímicas y numero de esporas de hongos micorrizico arbusculares en cinco estado sucesionales de un bosque altoandino en Guasca, Cundinamarca.
Asociación |
N° esporas/g |
Resistencia |
Temperatura del suelo |
pH |
M.O |
P |
Ca |
Mg |
K |
Fe |
Zn |
B |
|
|
Kg/cm² |
°C |
|
% |
ppm |
||||||
I |
15a |
2a |
11a |
4,5a |
4,7b |
5a |
0,5a |
0,2a |
0,2a |
442a |
2,7a |
0,2a |
II |
4a |
1,6ab |
11a |
4,1a |
10,7ab |
6a |
0,7a |
0,2a |
0,1a |
655a |
2,5a |
0,1a |
III |
12a |
1,4ab |
12a |
4,9a |
14,2ab |
3a |
0,5a |
0,1a |
0,1a |
503a |
2,2a |
0,2a |
IV |
15a |
0,7b |
12a |
4,2a |
16,6a |
4a |
0,6a |
0,3a |
0,2a |
561a |
2,6a |
0,2a |
V |
20a |
0,6b |
13a |
4,2a |
18,6a |
4a |
0,5a |
0,2a |
0,2a |
542a |
1,8a |
0,1a |
Letras diferentes en la misma columna representan diferencias significativas de la variable entre asociaciones vegetales por el test t (LSD) (p < 0,05).
Fuente de: los autores
Presencia de esporas de HMA
Se encontró que el estado sucesional con mayor número promedio de esporas fue el estado maduro (V) en el cual se registraron 20 esporas*g-1, entre tanto, los estados I y IV que corresponden a pionero e intermedio tardío, obtuvieron un valor promedio inferior, con 15 esporas*g-1. Finalmente el estado sucesional intermedio temprano (II) resultó tener el menor número de esporas (4 esporas*g-1). El análisis de varianza del número de esporas encontrados, muestra que no hay diferencias significativas (p < 0,05) entre la abundancia de esporas de HMA y los estados sucesionales del bosque evaluado (Tabla 2.) no obstante, se observa que hay mayor número de esporas en zonas de bosque maduro, donde el desarrollo estructural es mayor con respecto a los demás estados sucesionales; dicho resultado puede relacionarse con lo mencionado por Van der Heijden et al. (1998) quienes afirman que es posible que los HMA lleguen a potenciar la diversidad y productividad de una comunidad vegetal, dado que con una mayor diversidad y abundancia de esporas de HMA, la tendencia natural es que la vegetación que allí se sustenta, alcance un grado de complejidad igualmente mayor. Así mismo, Kernaghan (2005) afirma que existe una reciprocidad entre las comunidades de hongos formadores de micorrizas y las comunidades vegetales, esto es, que en la medida en que se da la asociación simbiótica, la composición de la comunidad vegetal establece un mayor grado de complejidad, aumentando el número de plantas hospederas disponibles para el hongo, lo que a su vez aumenta la productividad y estructura mejor la comunidad de HMA presente en la rizósfera de dicho ecosistema.
E
Cabe resaltar que para el caso del estado sucesional pionero (I), se observa, en contraposición a lo mencionado previamente, un alto número de esporas por gramo; se puede atribuir este resultado a la alta dependencia micorrizal que presenta este grupo de especies, del orden del 90% (Zángaro et al., 2000); y que responde a su necesidad de sobrevivir a fin de garantizar la continuidad de la sucesión. La simbiosis con el HMA se constituye entonces, en una estrategia necesaria para especies pioneras, lo que las lleva a desarrollar mecanismos que estimulen la esporulación del hongo y posterior colonización, como la alta concentración y secreción de metabolitos secundarios (flavonoides y estrigolactonas) en sus raíces, ya que estos compuestos son claves en la señalización planta-organismo (Akiyama, 2007).
Análisis de características fisicoquímicas del suelo comparadas con abundancia de esporas de HMA
En lo referido a las condiciones fisicoquímicas del suelo, se observan bajos valores de compactación, siendo en todos los casos menor o igual a 2,0 Kg*cm-2, característica que junto con el alto contenido de materia orgánica registrado, clasifica a este suelo, según lo establecido por Wagner & Medina(1998), como un suelo agronómicamente adecuado, dado que la menor compactación representa una mayor cantidad de espacios porosos, permitiendo retener igualmente un mayor volumen de agua, así como realizar el intercambio de gases; así mismo, facilitan la conductividad del agua y propician un mejor ambiente para el desarrollo radicular y de microorganismos. Los valores de pH encontrados oscilan entre suelos muy fuertemente ácidos (4.5 – 5.0) y extremadamente ácidos (<4.5) según la clasificación señalada por el IGAC (1999), lo que de acuerdo con Pereira et al. (2007) corresponde a un rango apto para la adecuada actividad de los hongos micorrízico arbusculares y el normal desarrollo del sistema radicular. Las diferentes concentraciones de elementos minerales registradas para cada una de las muestras evaluadas, permiten señalar que son suelos nutricionalmente deficientes (Fassbender & Boernemiza, 1987).
De las características fisicoquímicas evaluadas, el análisis de varianza (Tabla 2) muestra que solo la materia orgánica y la compactación presentan diferencias significativas (p < 0,05) entre las diferentes asociaciones vegetales o estados sucesionales, mientras que el contenido de los demás elementos evaluados no arrojó diferencias estadísticas significativas. Fue posible observar también
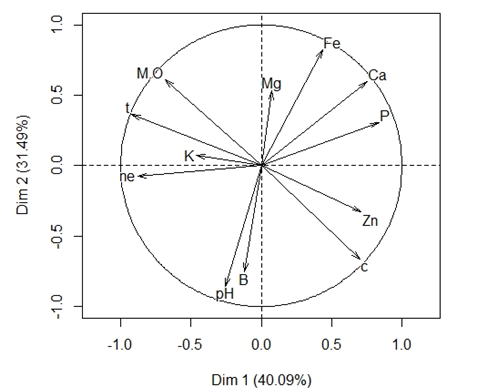
En la figura 1 se presentan los resultados del análisis de componentes principales (ACP) realizado con las variables de la tabla 2. Los primeros cuatro componentes fueron significativos (con valores propios mayores a 1); los dos primeros explicaron el 71,58% de la varianza total, mientras que los dos últimos explicaron solo el 28,42%.
Figura 1. Análisis de componentes principales de variables físico químicas del suelo en cinco estados sucesionales de bosque altoandino en Colombia. ne: número de esporas; M.O: Materia orgánica; t: Temperatura; K: Potasio; pH: potencial de hidrógeno; B: Boro; C: compactación; Zn: Zinc; P: Fósforo; Ca: Calcio; Mg: Magnesio; Fe: Hierro. Dim: Dimensión Fuente de: los autores
El gráfico de la figura 1 muestra que hay una relación positiva entre la presencia de materia orgánica y el número de esporas de HMA, mientras que esta relación es negativa con respecto a la compactación del suelo, esto se debe a que la presencia de materia orgánica disminuye la densidad aparente del suelo, es decir, aumenta la porosidad total, principalmente de los poros de mayor tamaño evitando los procesos de compactación (Quiroga et al., 2016) lo que resulta en una ventaja para el desarrollo del hongo micorrízico. Además de lo anterior se ha demostrado que en presencia de materia orgánica las hifas del HMA tienen facilidad de proliferación debido a que son áreas en descomposición de las cuales el hongo se puede beneficiar en términos de adquisición de nitrógeno (Hodge, 2017). Este resultado concuerda con lo reportado por Espitia & Pérez (2016) quienes evaluaron la diversidad de HMA en tres agroecosistemas con pastos, encontrando que los mayores valores de densidad de HMA se dan en suelos no compactados y por ende con mayores contenidos de matera orgánica.
El pH por su parte, a pesar de no presentar una variación alta, corrobora lo mencionado por (Safir & Duniway, 1991), quienes señalan que a medida que aumenta la acidez del suelo, disminuyen los niveles de solubilidad y disponibilidad en elementos como el fósforo, hierro y magnesio. Entre tanto, Peña-Venegas et al. (2007) afirman que un ligero incremento en el pH podría inducir la esporulación del HMA, debido a que disminuye el nivel de saturación del aluminio y mejora la capacidad de intercambio, lo que conlleva a una disminución del estrés de la comunidad microbiana y aumento en la eficiencia de sus procesos. Con lo anterior, se infiere que las características químicas del suelo presentan una alta sensibilidad a las variaciones de pH y que estas a su vez influencian la densidad de esporas en la rizósfera (Perez et al., 2011). Por otra parte, se observa que existe una relación negativa entre el contenido de zinc en el suelo, y el el número de esporas (Figura 1), similar a lo que sucede con el fósforo, esto puede deberse a que uno de los beneficios de la relación simbiótica HMA-planta es la de permitir a la planta la obtención de elementos insolubles como el fósforo y el zinc (Bucher, 2007), sin embargo, si hay una mayor concentración de estos elementos en forma iónica que sean asimilables por la planta, se limita la necesidad de la simbiosis disminuyendo la esporulación del hongo (Ping et al., 2014). Finalmente, la temperatura del suelo se ve relacionada positivamente con el número de esporas, en concordancia con lo señalado por Wild (1992) quien afirma que la actividad metabólica de los organismos aumenta a medida que la temperatura se eleva, por lo que favorece el desarrollo de los HMA y en general de la microbiota del suelo.
Análisis de vegetación
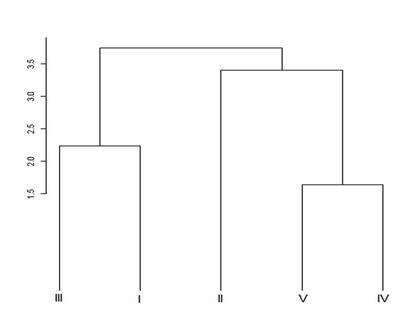
El análisis de agrupamiento de la figura 2. muestra que en dicho bosque las asociaciones vegetales están agrupadas en tres clados. En el primero de ellos se encuentran las asociaciones I y III, donde, para ambos casos, el género Weinmannia es dominante en la comunidad vegetal, y las características fisicoquímicas del suelo son semejantes, a excepción del contenido de materia orgánica, que es sustancialmente mayor en la asociación III, dicho comportamiento se debe a la presencia de individuos en estados de desarrollo más avanzados que aportan mayor cantidad de biomasa al suelo. A pesar de que difieren en el contenido de materia orgánica, las dos asociaciones presentan densidad de esporas similares (Tabla 2.), lo que puede relacionarse, como se mencionó antes, a la secreción de sustancias en la raíz de especies en estados tempranos de desarrollo, estimulando la esporulación del hongo. El segundo clado obtenido corresponde a las asociaciones IV y V, en donde se alcanzan los mayores valores de densidad de esporas, y a comparación con el primer clado, hay una estabilidad en las variables fisicoquímicas del suelo y en el desarrollo del bosque. Finalmente, la asociación II resulta ser diferente a las demás, presentando el menor número de esporas, se observa que en dicha asociación hay una acumulación de fósforo y de hierro que, sumado a las características estructurales, pueden estar limitando la esporulación del HMA.
Figura 2. Dendrograma de asociaciones vegetales presentes en cinco estados sucesionales de bosque altoandino Colombiano. I: Pionero (Weinmannio pinnatae - Miconietum theaezantis); II: Intermedio temprano (Ocoteo callophyllae - Weinmannietum pinnatae); III: Intermedio (Clusio multiflorae - Weinmannietum balbissianae); IV: Intermedio tardío (Rubo floribundi - Pteridietum aquilini); V: Maduro (Hedyosmo crenati - Myrcinetum coriaceae). Fuente de: Los autores
Conclusiones
Se registró presencia de esporas de hongos micorrízicos arbusculares en todos los estados sucesionales del bosque altoandino evaluado, sin encontrarse diferencias significativas de la abundancia de esporas entre dichos estados sucesionales; no obstante, se pudo observar que el bosque maduro es el que presenta mayor número de esporas y que hay una tendencia a disminuir en la medida que el grado de sucesión es menor, exceptuando lo observado en el estado pionero, que presentó alta abundancia de esporas. Por otra parte, la materia orgánica y la compactación del suelo, fueron las únicas características edáficas que presentaron diferencias significativas entre los estados sucesionales. El análisis de componentes principales dejó ver que el contenido de materia orgánica, la temperatura y el contenido de potasio son características que influenciaron positivamente la abundancia de esporas, así como el pH y el contenido de Boro aún cuando en menor proporción. Entre tanto, los altos contenidos de Hierro, Fósforo, Calcio y Zinc; así como un elevado índice de compactación, influenciaron negativamente la abundancia de esporas.
La comparación del número de esporas obtenidos con el contenido de materia orgánica en cada estado sucesional a excepción, nuevamente, del estado pionero, denotan una relación directamente proporcional, mientras que con la compactación del suelo, así como con el contenido de Fósforo, dicha relación es inversamente proporcional. Se concluye que el comportamiento diferencial observado para el estado sucesional pionero el cual presentó alto número de esporas en condiciones de bajo contenido de materia orgánica, alto contenido de Fósforo y alto índice de compactación, contrariando las tendencias observadas, pudo estar relacionado con la alta dependencia micorrizal que presentan las especies pioneras, llevándolas a desarrollar mecanismos para estimular la esporulación y el desarrollo del HMA.
Pese a no haber encontrado diferencias significativas, el número de esporas en los estados sucesionales deja ver que en zonas con mayor desarrollo estructural hay una abundancia de esporas más elevada y que puede disminuir dicha abundancia en la medida que disminuye su desarrollo estructural, sin embargo, hay otros factores como la fisiología de los grupos de especies presentes en cada estado sucesional, que pueden alterar dicho comportamiento, como se observó con el estado sucesional pionero.
Literatura citada