Alteraciones bioquímicas y metabólicas en el periodo de transición en vacas lecheras
Biochemical and metabolic disorders in the transition period
Rómulo Campos Gaona1
Adriana Correa-Orozco2
Gema Lucia Zambrano Burbano3
Andrea Ospina Cordoba4
1Médico Veterinario. Magister en Medicina Veterinaria. Doctor en Ciencias Veterinarias. Universidad Nacional de Colombia, sede Palmira, Colombia
2Médica Veterinaria Zootecnista. Magister en Ciencias Veterinarias. Candidata a Doctor en Ciencias Agrarias. Universidad Nacional de Colombia, sede Palmira, Colombia
3Zootecnista. Magíster en Ciencias Agrarias, Candidata a Doctor en Ciencias Agrarias. Universidad Nacional de Colombia, sede Palmira, Colombia
4Zootecnista. Estudiante de Maestría en Ciencias Agrarias. Universidad Nacional de Colombia, sede Palmira, Colombia
11rcamposg@unal.edu.co
2acorreao@unal.edu.co
3glzambranob@unal.edu.co
4paospinac@unal.edu.co
Resumen
La genética moderna ha logrado la selección de vacas con alta capacidad de producción de leche por lactancia, sin embargo, esta selección ha generado desajustes homeostáticos en el periodo de transición lo cual incide en la presentación de desórdenes metabólicos y fallas reproductivas, que repercuten negativamente en la rentabilidad de los sistemas lecheros. El periodo de transición constituye una etapa de profundos cambios metabólicos y endocrinos que generan un desafío fisiológico en el final de la gestación y el inicio de la lactancia. Para establecer estrategias que disminuyan la presentación o severidad de las alteraciones metabólicas y su impacto en el desempeño reproductivo, es necesario conocer los mecanismos bioquímicos que son afectados, principalmente sobre el metabolismo energético y lipídico que estimulan rutas compensatorias como la gluconeogénesis. El objetivo de la presente revisión fue describir las principales alteraciones bioquímicas que ocurren durante el periodo de transición en vacas lecheras, la ocurrencia de balance energético negativo y algunos síndromes metabólicos y cambios reproductivos que éstas presentan.Palabras clave: ácidos grasos no esterificados, cetosis, factor I del crecimiento similar a la insulina, gluconeogénesis, hígado graso.
Abstract
The modern genetic has achieved cows selection with high ability for milk production per lactation. However, this selection produced homeostatic imbalances in the transition period and this has led to metabolic disorders and reproductive failure, which negatively influence the profitability of dairy production. The transition period is a stage of profound metabolic and endocrine changes that produced a physiological challenge in the late pregnancy and early lactation. Understanding biochemical mechanisms that are affected, primarily on energetic and lipid metabolism, which stimulate compensatory routes such as gluconeogenesis, will allow to establish strategies to reduce presentation or severity of metabolic diseases and their impact on reproductive performance.
The aim of this review was to describe the mainly biochemical alterations that happen during the transition period in dairy cows, the occurrence of negative energy balance and some metabolic syndromes.
Key-words: nonesterified fatty acids, ketosis, insulin like growth factor I, gluconeogenesis, fatty liver.
Resumen Gráfico: Alteraciones Bioquímicas y Metabólicas en el Periodo de Transición en Vacas Lecheras
Introducción
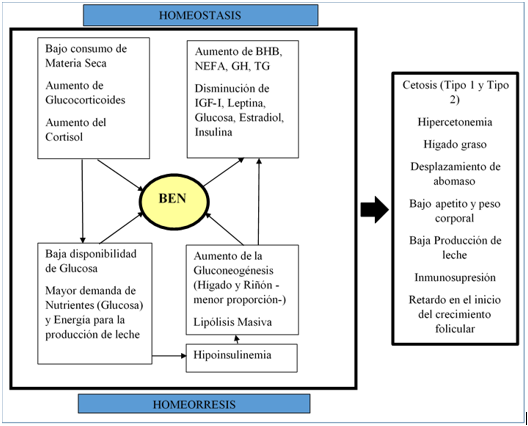
El periodo de transición en bovinos ha sido definido como la fase crítica en que la vaca próxima al parto, desde tres semanas antes al mismo hasta tres semanas después, cambia sus patrones metabólicos y se readapta para enfrentar las nuevas condiciones de lactancia; se considera es el periodo más sensible a las deficiencias energéticas (Markantonatos & Varga, 2017) por los profundos cambios endocrinos, metabólicos y homeostáticos que se dan en el animal, en el paso de un estado de preñez no lactante (vaca preparto) a un estado de no preñez y lactante (vaca postparto), los que constituyen un reto homeostático y homeorrético. Retomar las condiciones fisiológicas involucra una compleja integración de señales endocrinas y metabólicas centrales al eje somatotrófico que regula el fraccionamiento de energía, y al eje gonadotrófico que estimula la función reproductiva (Nicolini, 2014), los desafíos para atender dichas situaciones son rigurosos y generan una descompensación marcada en el animal, que no siempre éste logra regular y superar en forma adecuada, afectando su metabolismo y comportamiento productivo (Wang et al., 2017). En la lactancia, la vaca presenta el pico de producción entre los 30 a 60 días después del parto, simultáneamente el útero está en involución y el ovario retorna a la ciclicidad estral, lo cual puede implicar un conflicto entre estos procesos, particularmente si los cambios homeorréticos que ocurren en la lactancia temprana están desequilibrados y la vaca experimenta balance energético negativo (BEN), enfermedades metabólicas o ambos (Lucy, Butler, & Garverick, 2014).
Dado que la lactancia es ontogénicamente prioritaria, la síntesis láctea modificará la priorización metabólica de la energía, llevando progresivamente a la vaca a un BEN, del cual se derivarán en el corto plazo una serie de alteraciones metabólicas, como cetosis, hígado graso, desplazamiento de abomaso, entre otras (Overton, McArt, & Nydam, 2017), los cuales se agudizan en los animales considerados de alta producción (aquellos que usan como energía neta para lactancia, el equivalente al 70% de la energía total consumida). Además, esos drásticos cambios hormonales que se presentan durante el período de transición sumados a la depresión de consumo de materia seca, modifican sustancialmente el aporte de energía al animal, lo cual crea un alto riesgo de no lograr compensar la homeostasis energética y aumentar el BEN, lo cual se relaciona con la disminución en las concentraciones de estradiol, quien a su vez depende de mecanismos estereidogénicos para su síntesis (Cheong et al., 2016). Por consiguiente, la mayor demanda de nutrientes en el inicio de la lactancia, unida al menor ingreso de los mismos, llevan al animal a un BEN, que será el principal disturbio fisiológico en vacas lecheras (Baumgard, Collier, & Bauman, 2017).
Los mecanismos de obtención de energía en los rumiantes, dependen estrechamente de la fermentación bacteriana y la producción de ácidos grasos de cadena corta, en especial del ácido propiónico, como precursor principal en la síntesis de glucosa, imprescindible para el metabolismo energético celular de los diferentes tejidos (Aschenbach, Kristensen, Donkin, Hammon, & Penner, 2010). Por otra parte, la disminución en la concentración de glucosa, derivada del bajo consumo de materia seca durante el periodo de transición genera hipoinsulinemia, la cual conlleva a la movilización de energía, principalmente hacia la glándula mamaria para sintetizar lactosa – 72 g de glucosa por litro de leche producido- (Weaver & Hernandez, 2016), situación que promueve además la gluconeogénesis hepática (hasta 4 kg de glucosa al día) y actúa como un desencadenante de la lipolisis masiva (Leroy, Vanholder, Van Knegsel, Garcia-Ispierto, & Bols, 2008), ésta última para producir sustratos susceptibles de ser fuente energética a través de la beta-oxidación, en un intento metabólico de buscar fuentes energética alternas y corregir el BEN, dado que mientras se mantenga un BEN crítico (usualmente en las primeras tres semanas posparto), la respuesta tisular a la acción hormonal, favorecerá la lipolisis inhibiendo la lipogénesis, lo que causa aumento de ácidos grasos no esterificados (NEFA) y beta-hidroxibutirato (BHB) en sangre (Fiore et al., 2017). Se conoce que la alteración del metabolismo lipídico juega un papel importante en la patogénesis de muchas enfermedades en animales incluyendo cetosis, diabetes y resistencia insulínica (Gerspach et al., 2017; Overton et al., 2017). Así mismo, la función hepática es un punto central para el metabolismo y se torna especialmente crítica en el período de transición; durante esta etapa, una vaca lechera promedio moviliza 60 kg de grasa corporal y 20 a 25 kg de proteína corporal (Seymour, 2016).
Por otra parte, el estudio de las relaciones metabólicas, su regulación, componente genético y efectos hormonales (asociados al estrés y la disfunción ovárica), actualmente se desarrollan a través de nuevas concepciones fisiológicas y bioquímicas, consideradas mediante el uso de las técnicas llamadas “ómicas”, donde la metabolómica, genómica, proteómica entre otras han proporcionado nueva información para entender las interacciones biológicas que se presentan durante las fases críticas del periodo de transición (Vailati-Riboni, Elolimy, & Loor, 2016).
La presente revisión tuvo como objetivo analizar la asociación entre los mecanismos bioquímicos de ajuste homeostático durante el periodo de transición de vacas lecheras, la presentación de algunas alteraciones metabólicas y su incidencia en el balance energético negativo.
Depresión del consumo y período de transición
Los cambios adaptativos durante el período de transición, inician con la compensación metabólica por la disminución del consumo de alimento; como teorías que expliquen esta situación se ha postulado que el desequilibrio hormonal de los esteroides sexuales (mayor secreción de estrógenos y caída de la progesterona), unido a los cambios en el volumen y capacidad abdominal alterada al final de la gestación y a la oxidación de los nutrientes en el hígado, tanto exógenos como endógenos, además de la presencia de cortisol plasmático producto del estrés fisiológico del parto, contribuyen en diferente porcentaje a la caída del consumo y el inicio de la movilización grasa, respuesta compensatoria en la búsqueda de una fuente de energía alternativa que permita mantener la glicemia (Farman, Nandi, Girish Kumar, Tripathi, & Gupta, 2016). De la misma forma, se ha obtenido evidencia de la influencia endocrina (por ejemplo, leptina e insulina) sobre la fisiología del apetito y la respuesta metabólica asociada al mismo durante el periodo de transición (Nowroozi-Asl, Aarabi, & Rowshan-Ghasrodashti, 2016), la cual incide en los procesos bioquímicos, en especial sobre los ajustes energéticos.
Asimismo, cabe señalar que en bovinos en condiciones ecológicas de trópico, el consumo de alimento se ve afectado tanto por la calidad de los alimentos proporcionados, como por el efecto climático, donde la disipación de calor genera menor consumo y mayores requerimientos energéticos (Das et al., 2016). Los problemas de adaptación y depresión en la ingesta de alimento, pueden ser más severos en animales en crecimiento, como ocurre con las hembras de primer parto (Akins, 2016).
La principal consecuencia de la caída de la ingesta, que ocurre simultáneamente con el aumento en los requerimientos para mantener la lactancia, es la movilización severa de grasa, conocida como lipolisis y constituye uno de los principales procesos catabólicos, cuyo fin es suministrar sustratos para la gluconeogénesis y en conjunto estos cambios bioquímicos conllevan a la activación de procesos homeostásicos, no siempre exitosos (Kuhla, Metges, & Hammon, 2016).
Además, durante el BEN la insulina, el factor de crecimiento insulínico tipo I (IGF-I), la leptina y la glucosa presentan concentraciones bajas, mientras el BHB, los NEFA, la hormona del crecimiento (GH) y los glucocorticoides están elevados, estos y otros marcadores del metabolismo estrechamente relacionados con el hígado, pueden ser de amplia utilidad en el reconocimiento de rutas metabólicas de amplia actividad durante el periodo de transición (Butler, 2014; Galmessa, Kimothi, Mohammed, & Dang, 2017), que además influencian negativamente la fertilidad de forma indirecta, al modular el eje gonadotrópico y somatotrópico y de manera directa el ovario, folículo o ambiente uterino (Leroy et al., 2008).
Aumento de la gluconeogénesis
En los mamíferos superiores, la gluconeogénesis se asocia a la respuesta al ayuno y es fundamental en la homeostasis de la glicemia; aunque, la nueva glucosa sintetizada puede terminar incorporada al glucógeno hepático en ciertas condiciones postabsortivas. La gluconeogénesis como ruta metabólica anabólica permite la biosíntesis de glucosa a partir de precursores no glucémicos, en especial cuando éstos no se proveen en forma dinámica en el proceso digestivo; esta vía se produce mayoritariamente en el hígado y en menor grado en el riñón (Pérez-Mendoza, De Ita-Pérez, & Díaz-Muñoz, 2012). La gluconeogénesis se considera un proceso decisivo en el metabolismo energético de las vacas lecheras estabuladas, debido a que apoya la homeostasis de la glucosa, cuando el suministro de energía es insuficiente (Feijó et al., 2016; van Knegsel et al., 2014); igualmente, en animales en pastoreo favorece el metabolismo energético e incrementa la producción de leche, especialmente en vacas con limitantes en el aporte energético de carbohidratos solubles (Meléndez & Bartolome, 2017). Se considera que la glucosa es el sustrato energético más importante en el periodo de transición (Markantonatos & Varga, 2017); por consiguiente, durante la lactancia temprana en el rumiante, la gluconeogénesis es el principal mecanismo de obtención de energía, estimulada también por los elevados niveles de cortisol plasmático y basada principalmente en la utilización de propionato como sustrato (Ahmadi, Safi, Mortazavi, & Rokni, 2016).
Los ácidos grasos volátiles (AGV) provenientes de la fermentación ruminal de los carbohidratos, son la principal fuente de energía en los rumiantes y generan el 50% de la energía requerida por el animal, todos los carbohidratos que son suministrados en la dieta –forrajes o granos- serán fermentados, se absorberán y transportarán al hígado vía sistema portal enterohepático (Miller-Cushon & DeVries, 2017). En el hígado las vías metabólicas que toman los AGV son varias, en el caso del acetato la principal vía es la síntesis de acetilcoenzima A (acetil CoA) y por consiguiente capta productos provenientes de la beta-oxidación de ácidos grasos de cadena media y larga (Gómez & Campos, 2016). El propionato producido en el rumen es el precursor gluconeogénico primario en rumiantes, y la conversión de propionato en glucosa está controlada por la abundancia de la fosfoenolpiruvato citosólica (PEPCK-C), enzima fundamental para la gluconeogénesis hepática (Donkin, 2016; White et al., 2016). El mecanismo de control de esta ruta metabólica en bovinos es hormonal, ejercido por la insulina y el glucagón (Donkin, 2016); frente a la insulina cabe señalar que, las concentraciones de ésta pueden variar según la etapa de lactancia, la dieta, los desplazamientos prolongados y la disminución del consumo de materia seca, característico durante la preñez tardía y la lactancia temprana (Cohick, 2016).
Con el fin de aumentar la eficiencia energética, los rumiantes pueden aprovechar el lactato como fuente energética y la gluconeogénesis es una vía para controlar la acumulación de las concentraciones deletéreas de ácido D-láctico en plasma sanguíneo, debido a que cuando se absorbe en el rumen, éste isómero se metaboliza más lento que el L-isómero (Cuervo Vivas, 2017). Por otro lado, las enzimas microbianas en el rumen pueden aumentar la utilidad de ciertos sustratos glucogénicos derivados de la fermentación ruminal, que no son de fácil uso por los monogástricos (Dieho et al., 2017).
Ya en el hepatocito, el propionato se convierte a oxalacetato (OAA) a través de la propionil CoA carboxilasa mitocondrial (PCoAC), metilmalonil-CoA mutasa (MCM) y parte del ciclo de los ácidos tricarboxílicos (TCA); así mismo, el lactato y el principal aminoácido glucogénico alanina, se convierten inicialmente en piruvato en el citosol, antes de convertirsen en OAA por la piruvato carboxilasa mitocondrial (PC). El OAA puede ser entonces metabolizado por la fosfoenolpiruvato carboxicinasa (PEPCK) a fosfoenolpiruvato (PEP) y posteriormente a glucosa, o puede servir como aceptor de acetil CoA en el ciclo de los TCA (Donkin, 2016). De este modo, la estequiometría de la gluconeogénesis determina que la formación de PEP a partir de propionato, piruvato y algunos aminoácidos requiere la síntesis independiente de nicotinamida adenina dinucleótido (NADH) en el citosol para la posterior reducción de 1,3-difosfoglicerato en la gluconeogénesis (Mann et al., 2016).
Por otra parte, la alta prioridad metabólica para la gluconeogénesis conduce al drenaje de OAA del ciclo de los TCA (cataplerosis - salida del ciclo de los TCA). Por el contrario, el suministro de OAA al ciclo de los TCA mediante anaplerosis (entrada al ciclo de los TCA) sirve para reponer o compensar la pérdida de carbono debido a la cataplerosis (White, 2015). No obstante, aunque el OAA mitocondrial es el punto de fusión para la entrada de la mayoría de los sustratos lipídicos gluconeogénicos al ciclo de los TCA, éste no es anaplerótico y no puede estimular la oxidación de la acetil CoA (Gualdrón-Duarte & Allen, 2017). Lo anterior puede causar trastornos metabólicos, especialmente en la utilización de acetil CoA proveniente de la beta-oxidación, provocando cetosis y en algunos casos hígado graso (Ahmadi et al., 2016).
Metabolismo de los ácidos grasos no esterificados (NEFA) en el hígado
Durante los períodos de BEN, los triglicéridos (TG) del tejido adiposo son hidrolizados por las lipasas a glicerol y NEFA y son liberados a la circulación (Kuhla et al., 2016). En el hígado, los NEFA son transportados por la carnitina aciltransferasa 1 hacia la mitocondria por unión a la carnitina y entran a la beta-oxidación, en donde cada ciclo corta el ácido graso en moléculas de dos carbonos y produce una molécula de acetil CoA, una de flavin adenin dinucleótido reducido (FADH2) y una de NADH (Han van der Kolk, Gross, Gerber, & Bruckmaier, 2017). La beta-oxidación se mantiene hasta que se divida la cadena completa en unidades de acetil CoA y en el caso de un ácido graso de cadena impar, hasta que haya un propionil CoA (Han van der Kolk et al., 2017). Posteriormente, los NEFA (a partir de la acetil CoA) pueden tener cuatro destinos posibles: 1) oxidación completa en el ciclo de los TCA y producción de ATP (energía), H2O y CO2; 2) oxidación parcial a través de la cetogénesis (producción de cuerpos cetónicos acetona, ácido acetoacético y ácido betahidroxibutírico); 3) esterificación a TG para empaquetamiento en lipoproteínas de muy baja densidad (VLDL) y transporte fuera del hígado; o 4) almacenamiento de esos TG en el tejido hepático (Bisinotto et al., 2012; Sun et al., 2016).
Cuando las vacas movilizan tejido adiposo se puede exceder la capacidad del ciclo de los TCA para oxidar completamente la acetil CoA (White, 2015), lo que conlleva a un incremento en la producción de cuerpos cetónicos (Han van der Kolk et al., 2017; White, 2015) y deposición de TG en el tejido hepático, desencadenando el inicio del síndrome de cetosis e hígado graso, respectivamente (White, 2015; Zarrin, Grossen-Rösti, Bruckmaier, & Gross, 2017). Esa deposición de TG se debe a que el hígado bovino tiene una capacidad limitada para sintetizar y secretar VLDL, por lo tanto, se compromete la salida de TG durante los periodos de gran captación hepática de NEFA (Bisinotto et al., 2012). Aunque el mecanismo hepático de hígado graso bovino y cetosis no está completamente entendido, es claro que la regulación de la absorción, lipólisis, oxidación y almacenamiento de TG juega un papel clave en el inicio y evolución de estos desórdenes (Han van der Kolk et al., 2017).
La capacidad del hígado para oxidar acetil CoA está determinada por el total del intermediarios del ciclo de los TCA, los cuales a su vez son afectados por el balance entre sus tasas de entrada (anaplerosis) y salida (cataplerosis) y su flujo a través del ciclo (determinado por la concentración y actividad enzimática y el estado redox) (Han van der Kolk et al., 2017; White, 2015). Los intermediarios del ciclo de los TCA son reemplazados continuamente, por ejemplo, el succinil CoA a partir del propionato y el oxalacetato (OAA) vía piruvato desde el lactato. Otros intermediarios del ciclo se pueden reponer por la descarboxilación de diferentes aminoácidos gluconeogénicos (alanina, treonina, serina, cisteína, valina, isoleucina, metionina, entre otros) (Gualdrón-Duarte & Allen, 2017).
Para el caso del OAA, el cual sirve como un transportador de carbonos para el ciclo de los TCA (White, 2015), este puede ser metabolizado a fosfoenolpiruvato y de ahí a glucosa vía gluconeogénesis (salida del ciclo de los TCA - cataplerosis) o puede metabolizarse en el ciclo de los TCA y servir como un aceptor de acetil CoA (Donkin, 2016). Por otra parte, la carboxilación de piruvato a OAA incrementa inmediatamente la reserva del OAA (entrada al ciclo de los TCA – anaplerosis) (White, 2015). Asimismo, los carbonos del propionato también pueden proveer OAA adicional si no son sacados de ciclo de los TCA hacia la gluconeogénesis (White, 2015). Por tanto, cuando hay una mayor gluconeogénesis (por ejemplo, durante el pico de lactancia) se hace uso de las reservas de OAA, lo cual reduce su disponibilidad para aceptar acetil CoA en las mitocondrias de los hepatocitos y en consecuencia se limita la oxidación completa de los ácidos grasos a través del ciclo de los TCA, y esta acumulación de acetil CoA conlleva a la activación y aumento de una vía metabólica alternativa, la cetogénesis (Bruckmaier & Gross, 2017). En conclusión, el suministro de OAA al ciclo de los TCA mediante la reacción anaplerótica sirve para reponer o compensar la pérdida de carbono debido a la cataplerosis, lo cual indica que el balance entre cataplerosis y anaplerosis es crítico específicamente en el periodo de transición, cuando la demanda de glucosa y energía superan la disponibilidad (Gualdrón-Duarte & Allen, 2017; White, 2015).
Es necesario señalar que la producción de NEFA y cuerpos cetónicos (por ejemplo, BHB) que actúan como fuente de energía alternativa son una secuela normal de la adaptación al BEN, y así, ciertas concentraciones circulantes de esos metabolitos energéticos en la sangre (< 1,2 mmol/L de BHB) son normales durante el inicio de la lactancia (Ruoff, Borchardt, & Heuwieser, 2017). Sin embargo, una excesiva producción de NEFA y BHB son indicativos de una pobre respuesta adaptativa al BEN (Ruoff et al., 2017; Zarrin et al., 2017) y están asociados con detrimento de los resultados productivos y de salud (McArt, Nydam, Oetzel, Overton, & Ospina, 2013), ya que generan patologías como cetosis o hígado graso (White, 2015).
Exceso de movilización grasa y presentación de cetosis
La cetosis corresponde a una acumulación de cuerpos cetónicos en la sangre cuando se movilizan grandes cantidades de grasa corporal y no hay suficientes carbohidratos para facilitar la beta oxidación (Han van der Kolk et al., 2017). Sin embargo, se conocen dos tipos de cetosis: tipo I y II. La cetosis tipo I ocurre cuando la demanda de glucosa supera la capacidad del hígado para la gluconeogénesis, con las vías gluconeogénicas que podrían estar en el punto más alto de estimulación, pero el suministro de precursores de glucosa es insuficiente para permitir la máxima producción de glucosa (Sundrum, 2015). En otras palabras, la producción de glucosa está limitada por el suministro de sustrato (Sundrum, 2015) o los procesos productivos están descompensados por la densidad y aporte energético de la ración (Wang et al., 2017). Desde el punto de vista de la práctica clínica, la cetosis tipo I es el resultado de la insuficiencia de energía metabolizable en relación con las demandas fisiológicas, y ocurre típicamente entre la semana 3 y 8 posparto por el aumento en la producción de leche y disminución del consumo de materia seca, lo que crea un "vacío de energía" (Cooper, 2014); llama la atención que la insulina en la sangre es normalmente baja en estos animales, comparable a los diabéticos humanos tipo 1 (Cooper, 2014).
Por otra parte, la cetosis tipo II se produce cuando se entregan grandes cantidades de NEFA al hígado, pero la gluconeogénesis y la cetogénesis no están en el punto más alto de estimulación (Sundrum, 2015). La captación mitocondrial de NEFA no es tan activa como en la cetosis tipo I, por lo cual los NEFA que no son utilizados para la síntesis de cuerpos cetónicos son esterificados a TG en el citosol y como la capacidad de transporte a otros tejidos mediante VLDL puede ser limitada se acumulan TG en el hígado (Han van der Kolk et al., 2017; Sundrum, 2015). El hígado graso y la cetosis tipo II generalmente se ven en forma clínica poco después del parto (una a dos semanas), mucho antes de la producción máxima de leche (Sundrum, 2015). En campo, ésta forma de cetosis está asociada a una excesiva movilización de grasa y es más común en vacas de alto mérito genético y sobrecondicionadas (condición corporal superior a 3,5 en la escala 1-5 ya genera riesgo) o en granjas con pobres sistemas de manejo y alimentación de la vaca de transición (Cooper, 2014). Además, se caracteriza por concentraciones de insulina normales a altas y resistencia periférica a la misma, lo cual la hace comparable con la diabetes tipo II en humanos (Cooper, 2014).
Finalmente, la hipercetonemia marcada se manifiesta clínicamente con un apetito reducido, pérdida rápida de peso y disminución en la producción de leche (Marutsova, Binev, & Marutsov, 2015; Pryce et al., 2016) y, además, puede generar inmunosupresión, la cual se asocia con el aumento de patologías infecciosas (Lacasse, Vanacker, Ollier, & Ster, 2017).
Lipomovilización incompleta en hígado y riesgo de lipidosis hepática
Según Pérez-Mendoza et al. (2012), el hígado es el órgano central del metabolismo y aunque su estructura celular parece ser homogénea, diversos estudios histoquímicos y bioquímicos han encontrado diferencias entre los hepatocitos, tanto en la presencia como en la concentración de diversas enzimas implicadas en varias vías metabólicas. A este fenómeno se le ha denominado zonación metabólica o heterogeneidad funcional, que es afectada por situaciones como la lipomovilización y la alteración de sus depósitos de grasa.
Por lo tanto, al considerar el hígado un órgano vital en la homeostasia lipídica de todo el organismo, es importante conocer los cambios en la composición energética y grasa en la dieta de los rumiantes durante el período de transición, lo cual es crítico para la regulación fisiológica (Contreras, Strieder-Barboza, & Raphael, 2017). Precisamente en éste periodo, el examen histopatológico evidencia un grado de moderado a severo de acumulación intracelular grasa en el hígado, especialmente en vacas con cetosis (Djokovic et al., 2013). Las lipasas hidrolizan varios tipos de lípidos incluyendo TG, diglicéridos, monoglicéridos, y ésteres de colesterol, tras la activación por la proteína quinasa A (PKA) (Contreras et al., 2017). El incremento de TG en el hígado, disminuye el aporte de estos compuestos a lípidos estructurales (colesterol libre, esteres de colesterol, fosfolípidos); los precursores de energía como citrato ocasionan un menor acúmulo de reservas de energía por la menor síntesis de glicógeno, con lo cual aumenta el riesgo de alteraciones metabólicas hepáticas que deben ser monitoreadas, antes de que la acumulación grasa sea irreversible (Cooper, Green, Macrae, Hayton, & Patton, 2011).
La acelerada lipomovilización que surge principalmente ante un BEN severo, algunas veces más evidente en animales que llegan al preparto con excesos de acúmulo de grasa, conduce a alteraciones hepáticas, que podrían definirse como disminución de la función y acumulación de metabolitos grasos al interior del hepatocito, ésta última con diferentes niveles de depósito y alteración funcional; la condición está estrechamente relacionada con los mecanismos gluconeogénicos y los niveles plasmáticos de insulina (Donkin, 2016).
Dependiendo del grado de infiltración de lípidos en el hepatocito, se han considerado cuatro grandes categorías que van desde leve a severa, ésta última con acumulación superior al 75%; sin embargo, el grado de infiltración y su sentido fisiológico es difícil de definir (Cooper et al., 2011). La pérdida de la arquitectura celular, afecta el funcionamiento y permite que aparezcan alteraciones bioquímicas que pueden ser identificadas por los cambios en los indicadores metabólicos y enzimáticos con los cuales se monitorea la función hepática (Ceciliani, Lecchi, Urh, & Sauerwein, 2017; González, Muiño, Pereira, Campos, & Benedito, 2011). Finalmente, el hígado graso es considerado una de las enfermedades metabólicas más severas en vacas lecheras y como se mencionó antes, normalmente ocurre como secuela de la cetosis tipo II (Cooper, 2014; Djokovic et al., 2013).
De otra parte, la concentración de proteínas hepáticas es fundamental para evitar la acumulación de lípidos en el hígado, la alteración de su oxidación o su velocidad de exportación del tejido (McFadden, 2017). La proteína quinasa C (PKC) es una familia de enzimas asociadas a las proteínas quinasas que se encuentra en hepatocitos y adipocitos, junto con la apoliproteína B, regula la secreción de VLDL y el receptor de lipoproteínas de baja densidad (LDLR), los cuales son responsables de la salida de los lípidos desde el hígado hacia el torrente circulatorio y la recolección de colesterol libre (FC) por lipoproteínas de alta densidad (HDL) (Ahmadi et al., 2016). La PKC, asociada a la modificación postransduccional de la apoliproteína A-I, induce la activación de la lecitina-colesterol acil transferasa y, la carnitina palmitoil transferasa en la mitocondria que participa en las reacciones de beta-oxidación y cetosis.
A través del tiempo, se han empleado diferentes opciones de diagnóstico de hígado graso, siendo actualmente el monitoreo por ultrasonografía (Fiore et al., 2017) la técnica más aceptada, descartando la invasiva biopsia hepática (Djokovic et al., 2013), tornándose más concluyente que los análisis de patología clínica, cuyos resultados pueden ser imprecisos (Cooper et al., 2011); sin embargo, en los últimos años se ha empleado en forma prometedora las proteínas de fase aguda (haptoglobina, proteína C reactiva) y enzimas como la paraoxonasa, de utilidad diagnóstica dada su alta correlación con los niveles de NEFA, VLDL y TG (Overton et al., 2017).
Alteración hepática y disminución de la síntesis del factor de crecimiento insulínico Tipo I (IGF-I)
El eje somatotrófico, que está constituido por la GH, IGF-I, sus proteínas de unión (IGFBPs) y sus receptores (Lucy, 2012), coordina las adaptaciones fisiológicas para preservar la homeostasis metabólica durante los estados catabólicos (desnutrición y parto) (Kuhla et al., 2016). Este eje está involucrado como una vía metabólica clave que facilita la transición desde el final de la preñez al inicio de la lactancia (Farman et al., 2016). Se conoce que en el posparto temprano ocurre un desacople hormonal del eje debido a que durante el inicio de la lactancia disminuyen las concentraciones sanguíneas de glucosa e insulina (Lucy et al., 2014), esta última responsable por la baja regulación del receptor de la GH específico del hígado tipo IA (GHRIA) (Oliveira et al., 2016), lo que resulta en altas concentraciones sanguíneas de GH y bajas de IGF-I (Li et al., 2016), además asociadas a la reducción en la expresión del ARNm de IGF-I en los hepatocitos (Li et al., 2016). Dado que la insulina controla la síntesis y secreción del IGF-I hepático, las concentraciones de glucosa, insulina e IGF-I están correlacionadas (Berry, Friggens, Lucy, & Roche, 2016).
El IGF-I es una hormona peptídica, con funciones endocrinas, paracrinas y autocrinas (Livingstone, 2013), sintetizada y secretada en su mayor parte por el hígado (80%), mientras el restante (20%) es sintetizado por lo general en células del tejido conectivo, en donde regula la supervivencia celular (Takada, Takada, & Fujita, 2017). En su transporte por la circulación sanguínea se une a una de sus seis proteínas transportadoras (IGFBPs1-6), especialmente la IGFBP3 (Takada et al., 2017). La disponibilidad del IGF-I está estrechamente regulada por las IGFBPs y la subunidad ácido lábil (ALS), las cuales aumentan su vida útil y sirven como medio de transporte a tejidos específicos (Aguirre, Rodríguez De Ita, de la Garza, & Castilla-Cortazar, 2016). Los IGFs circulantes están en su mayoría unidos a complejos ternarios que consisten en IGF-I o IGF-II, IGFBP-3 y ALS, los cuales tienen una vida media relativamente larga en la circulación (10-16 h), por lo cual actúan como una reserva endocrina de IGFs (Livingstone, 2013). También puede haber IGFs libres o en complejos binarios, estos últimos con acceso a los tejidos y pueden actuar como un reservorio pericelular, a diferencia de los complejos ternarios, que son incapaces de cruzar el endotelio capilar (Livingstone, 2013). Para el caso de IGF-I, el tamaño de la reserva está determinado por la cantidad de IGFBP-3 y ALS circulante (Livingstone, 2013).
Aunque las concentraciones sistémicas de IGF-I podrían explicarse por una reducción correspondiente en el ARNm hepático, es evidente que la expresión alterada de proteínas de unión moduladora y receptores asociados con la señalización paracrina también puede contribuir a la hipótesis de actividad de IGF-I suprimida (Fenwick et al., 2008). La cantidad significativamente reducida de expresión hepática de ARNm, de ALS hepática y otros componentes ternarios (IGFBP-3, -5) en vacas con BEN severo en comparación con las vacas con BEN leve indica que un efecto principal del BEN puede ser evitar o al menos atenuar la entrega y la estabilidad endocrina de IGF (Farman et al., 2016; Fenwick et al., 2008). Igualmente, el efecto de la reducción del receptor de GH (GHR) total en vacas con BEN severo, implica una señalización reducida de GHR y una síntesis atenuada de IGF-I (Fenwick et al., 2008). Además, en vacas en el último mes de gestación con concentraciones plasmáticas totales de IGF-I bajas, estas se pueden atribuir a una menor producción hepática de ALS y una expresión de ARNm de IGFBP-4 más alta, esta última proteína responsable de inhibir la acción de IGF-I en el nivel del tejido y por lo tanto puede también afectar negativamente la biodisponibilidad de éste (Piechotta et al., 2013). El escenario general apunta a un ambiente endocrino de síntesis, estabilidad y biodisponibilidad de IGF-I reducida que posiblemente inclina la balanza a favor de IGF-II (Fenwick et al., 2008).
Por otra parte, también se ha propuesto que el IGF-I afecta el ensamblaje de las VLDL, lo cual sería un factor importante en el desarrollo del hígado graso (McFadden, 2017). La apolipoproteína (Apo) B100, Apo E, la proteína de transferencia de triglicéridos microsomal (MTTP) y el LDLR son los componentes de proteínas estructurales y reguladoras más importantes para el ensamblaje de VLDL (Li et al., 2016). En cultivo in vitro de hepatocitos de vaca, el IGF-I aumenta la abundancia de expresión de ApoB100, ApoE, MTTP y LDLR y como resultado, también aumenta la concentración de VLDL, lo cual indica que posiblemente la alteración en la síntesis de IGF-I afecta el ensamblaje de VLDL y luego participa en el desarrollo del hígado graso en las vacas lecheras (Li et al., 2016).
A su vez el impacto negativo de la lipidosis hepática sobre la reproducción se explica parcialmente por la menor síntesis de IGF-I en los hepatocitos, lo cual retarda el inicio del crecimiento folicular en el posparto temprano, y por la disminución en la respuesta inmunológica, como efecto de la menor secreción de proteínas en el hígado, presentando inmunodepresión asociada a retardo en la involución uterina y metritis (Cohick, 2016; Oliveira et al., 2016). Por lo tanto, el IGF-I endocrino es indicativo de la competencia metabólica para el restablecimiento de la fertilidad durante el período postparto en vacas con BEN (Cohick, 2016), debido a que tiene efectos directos sobre el hipotálamo, la hipófisis y el ovario, y en este último estimula la proliferación de células de la granulosa, promueve la esteroidogénesis, la foliculogénesis, ovulación, fertilización, implantación y el desarrollo embrionario (Rivas, Suárez, & Ramírez, 2011). Por consiguiente, es factible que el papel que juega el IGF-I en la partición de nutrientes, como una señal del estado bioenergético, represente un mecanismo clave que relacione el estado nutricional con el eje reproductivo (Butler, 2014).
Conclusiones
Se presentan los principales mecanismos bioquímicos durante el período de transición, en especial los relacionados con el metabolismo energético y la incidencia de éste en los procesos de homeostasis, la presentación de algunos desórdenes metabólicos y en la activación del eje gonadotrópico en vacas con marcada selección a la producción lechera.
Los ajustes en las rutas bioquímicas del metabolismo energético de origen hepático, que oxidan y sintetizan los nutrientes le permiten a la vaca lechera enfrentar los altos requerimientos energéticos durante el periodo de transición, mediante el aumento de la gluconeogénesis y la beta oxidación. Sin embargo, cuando hay un desequilibrio entre estas dos rutas, principalmente por un exceso en la lipolisis o por la falta de sustratos gluconeogénicos, se presentan alteraciones metabólicas como cetosis e hígado graso. Así mismo, los cambios generados por el BEN tienen impacto en el hígado, afectando la síntesis de proteínas como el IGF-I, el cual a su vez tiene efectos en el eje reproductivo.
Literatura citada