Liberación ruminal de cuatro fuentes inorgánicas de magnesio por medio de la técnica in situ en ganado holstein en trópico alto
Release ruminal of four inorganic sources of magnesium through the technique in situ in holstein cattle in high tropic
libertação no rúmen de quatro fontes inorgânicas de magnésio pela técnica in situ em Holsteins elevados trópicos
Wilmer Alfonso Cuervo Vivas1 & Héctor Jairo Correa Cardona2
1Zootecnista, Especialista en Nutrición Animal, Magister en Ciencias Agrarias. 2Zootecnista, Magister en Nutrición Animal, Doctor en Ciencias de la Producción Animal.
1Especialización Nutrición Animal Sostenible. Escuela de Ciencias Agrícolas, Pecuarias y del Medio Ambiente -ECAPMA, Universidad Nacional Abierta y a Distancia –UNAD. Bogotá, Colombia. 2Docente Asociado Departamento de Producción Animal,
Universidad Nacional de Colombia, sede Medellín. Colombia.
1wilmer.cuervo@unad.edu.co, 2hjcorreac@unal.edu.co
Resumen
El Magnesio, abundante en praderas pastoreadas por bovinos de leche, actúa como un cofactor en reacciones enzimáticas vitales para las principales vías metabólicas y presenta desaparición ruminal elevada (87%) aunque se desconoce su liberación en fuentes inorgánicas. Con el objetivo de evaluar su solubilización ruminal se incubaron cuatro fuentes inorgánicas (óxido de Mg (MgO), sulfato de Mg (MgSO4), cloruro de Mg (MgCl2) y carbonato de Mg (MgCO3)) determinando su desaparición de la materia seca (DMS) en 3 vacas Holstein (vacías, no lactantes) canuladas al rumen (620 ± 14 kg y 7 años) en el municipio de Santa Elena, Antioquia, Colombia (2538 msnm; bH – MB; 16ºc) consumiendo kikuyo (Cenchrus clandestinus (Hoechst Ex Chiov) Morrone) (Chemisquy et al 2010). Por triplicado se incubaron 0,5 g de cada fuente en bolsas de nailon con dos tamaños de poro (25 y 50μm), en el rumen de las vacas para evaluar DMS en tres tiempos (0, 12 y 24h). Se utilizó un diseño en bloques completos al azar con un arreglo factorial 2x3x4 (2 tamaños de poro, 3 tiempos de incubación y 4 fuentes minerales) y prueba de Tukey para significancia estadística de las diferencias. La incubación ruminal debe realizarse con tamaño de poro no superior a 50um, siendo MgCl2 y MgSO4 mas solubles en rumen, mientras que óxidos y carbonatos presentan solubilidad baja o nula dentro de las primeras 24 horas, lo cual puede dar luces para su utilización como suplemento mineral dentro de la dieta solida o agua de bebida del ganado lechero.
Palabras claves: holstein, líquido ruminal, tamaño de poro, kikuyo, ANKOM, BRIONI.
Abstract
Magnesium, abundant in grasslands grazed by cattle of milk, acts as a cofactor in enzymatic reactions vital to major metabolic pathways and presents disappearance ruminal high (87%) Although his release from inorganic sources is unknown. With the objective of evaluating their solubilization ruminal incubated four inorganic sources (Mg oxide (MgO), Mg sulfate (MgSO4), Mg chloride (MgCl2) and Mg carbonate (MgCO3)) determining its disappearance of dry matter (DMS) in 3 Holstein cows (empty, not nursing) placed in cannula to the rumen (620 ± 14 kg and 7 years) in the municipality of Santa Elena, Antioquia , Colombia (2538 masl; bH - MB; 16 ºc) eating Kikuyu (Cenchrus clandestinus (Hoechst Ex Chiov) Morrone) (Chemisquy et al 2010). 0.5 g of each source in two pore sizes nylon bags and incubated in triplicate (25 and 50μm), in the rumen of cows to evaluate DMS in three times (0, 12, and 24 h). A design was used in the randomized complete block factorial 2 x 3 x 4 (2 pore sizes), 3 days of incubation and 4 mineral sources and Tukey test for statistical significance of the differences. The ruminal incubation must be done with no more than 50um, MgCl2 and MgSO4 pore size more soluble in rumen, while oxides and carbonates are low or no solubility within 24 hours, which can give light for use as a mineral supplement within the solid diet or dairy cattle drinking water.
Key-words: Holstein, rumen fluid, pore size, kikuyus, ANKOM, BRIONI.
Introducción
El ion metálico Magnesio Mg es un catión intracelular que actúa como coenzima para reacciones enzimáticas vitales en las principales vías del metabolismo energético (NRC, 2001), su deficiencia es poco común en la mayoría de los animales domésticos (Aikawa, 1981, Underwood & Suttle, 1999). No obstante, cuando se ha observado bajas sensibles en el contenido de Mg en la dieta, se ha relacionado con el exceso de fósforo y potasio en la misma (Schonewille et al, 2004), observándose en animales malformaciones en las extremidades e incluso mortalidades, particularmente en especies como la aviar y la porcina (Atteh & Leeson, 1983; 1986).
El bajo contenido de Mg (Tomas & Potter, 1976; Field & Munro, 1977) y el excesivo contenido de potasio en forrajes altamente fertilizados (Schonewille et al., 1998; 2007) se ha relacionado con altas incidencias de hipomagnesemia. Los forrajes tropicales presentan un contenido de Mg que oscila entre 0,36% para gramíneas y 0,28 % para leguminosas (Minson, 1990), mientras que los alimentos concentrados (energéticos y proteicos) presentan un mayor contenido de Mg que los forrajes (Adediji & Suttle, 1999). El bajo contenido de Mg en forrajes y en cereales, menos frecuente, ha sido identificado como la principal causa de hipomagnesemia (Tomas & Potter, 1976; Field & Munro, 1977). Este tipo de desórdenes metabólicos, sin embargo, son comunes en zonas templadas y específicamente con rebrotes de forraje en la primavera, mientras que en regiones tropicales son poco frecuentes (Littledike et al., 1983). El elevado contenido de K en forrajes típicos del trópico alto como el kikuyo -hasta 5,12% de la MS- (Correa et al., 2008) están relacionados con disfunciones en el metabolismo del Ca y del Mg (Goff, 2006).
En los sistemas de producción ganadero, el principal, y en ocasiones el único, suplemento dietario es la sal mineralizada (MADR, 2010) y en ganado en periodo preparto, se han utilizado mezclas de macrominerales en forma de sales anionicas combinándolas con suplementación de Ca y Mg para la prevención de la hipocalcemia e hipomagnesemia (Underwood & Suttle, 1999, Drackley et al., 2005). El contenido de Mg de estas fuentes inorgánicas va desde 5% en el fosfato de Mg hasta un 52% en la magnesita calcinada (FEDNA 2012). En este sentido, sales de Mg como el carbonato, cloruro, oxido y sulfato han presentado valores de biodisponibilidad altos (McDowell 1996), su absorción oscila entre 30 y 70% (NRC 2001) y su solubilidad en medios acuosos depende en gran medida al tipo de sal (Horn & Smith 1978., NIST 2011).
La técnica In situ busca incubar diferentes materiales -nutricionales y no nutricionales- en bovinos, ovinos y caprinos fistulados y equipados con una cánula. El rumen es el principal sitio de absorción del Mg en rumiantes adultos (Martens & Blume, 1986; Care et al., 1984; Greene et al., 1983, Church 1988), sin embargo, un pH bajo (Cook et al. 1994) sumado a consumo de forrajes con altos contenidos de K (Fontenot et al, 1973, Tomas & Potter 1976) pueden disminuir su solubilidad. Se han reportado, valores iníciales de liberación ruminal de Mg en pasto kikuyo superiores al 70% (Correa et al. 2008) y hasta 93% a las 72 horas de incubación en rumen (Moseley & Griffiths 1984, Van Eys & Reid 1987) No obstante, Se desconocen trabajos en los que se haya estimado la liberación ruminal del Mg a partir de fuentes Inorgánicas incubadas como sales puras y utilizando la técnica In situ. Por ello el objetivo de este trabajo fue evaluar la desaparición de cuatro fuentes inorgánicas de Mg por medio de la técnica in situ utilizando dos tamaños de poro en la bolsa de nailon.
Materiales y métodos
Localización
Este estudio se realizó en la hacienda Paysandú de la Universidad Nacional de Colombia, ubicada en el municipio de Santa Elena, Antioquia, Colombia, a 2538 msnm y que pertenece a la zona de vida de Bosque húmedo montano bajo (bH – MB) (Holdridge, 1996).
Animales experimentales
Para la investigación se incluyeron tres vacas de la raza Holstein, adultas no lactantes, canuladas al rumen, con un peso y edad promedio de 620 ± 14 kg y 7 años, respectivamente. Dichos animales fueron alimentados a voluntad con pasto kikuyo. Las vacas experimentales fueron trasladadas desde el potrero hacia un corral de manejo a las 07:00 am, donde se realizó la extracción de la tapa de la cánula y la incubación de los sustratos a evaluar.
Preparación de las muestras
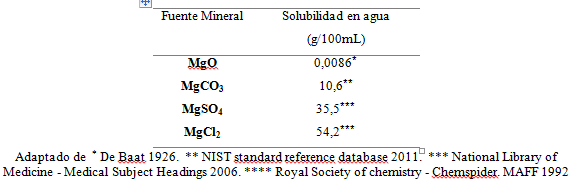
Se utilizaron 0,5 g de cada una de las fuentes inorgánicas a evaluar; oxido de Mg (MgO), sulfato de Mg (MgSO4), cloruro de Mg (MgCl2) y carbonato de Mg (MgCO3) cuyas solubilidades se presentan en la Tabla 1. Dichos compuestos fueron empacados en dos tipos de bolsas de nailon: bolsas FNº 57 Ankom ® con un tamaño de poro de 25 μm y dimensiones de 5 x 4 cm (para obtener un área efectiva por bolsa de 37 cm2) compuestas por un material tipo poliéster, y bolsas elaboradas con tela nacional (Brioni®-Lafayette) con un tamaño de poro de 50µm, implicando un menor costo. Este proceso buscaba evaluar la factibilidad de incluir bolsas de tela nacional en lugar de las bolsas Ankom ® que son importadas y tienen un alto costo para los experimentos relacionados con la técnica in situ.
Tabla 1.
Solubilidad en medios acuosos de las fuentes minerales evaluadas
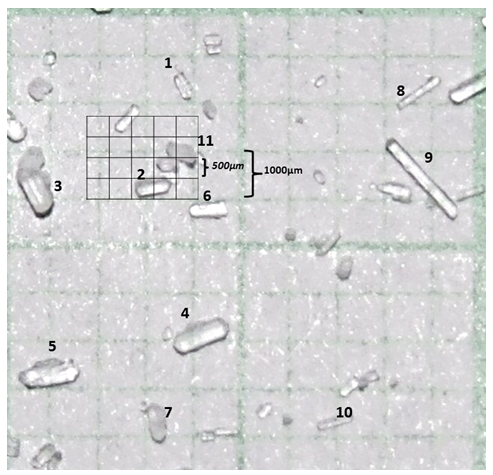
Para realizar la determinación del tamaño de partícula de cada fuente mineral, tomando fotografías con zoom óptico de 20X a las partículas de las cuatro fuentes minerales a evaluar, ubicándolas sobre una hoja de papel milimetrado con divisiones de 500 y 1000µm (ver Anexos), esto con la finalidad de establecer su relación con la DMS en cada tamaño del poro de las bolsas evaluadas. Se tomaron 11 observaciones de cada una de las fuentes minerales evaluadas y se obtuvo el promedio y la desviación estándar del tamaño de cada una de las fuentes. En cada una las bolsas evaluadas (Ankom ® y tela nacional), se depositaron 0,5 g de muestra (Goering & Van Soest, 1970) lo que corresponde a una relación tamaño de la muestra/superficie de la bolsa de 13.5 mg/cm2 basado en las recomendaciones publicadas por Van Hellen & Ellis (1977) para la técnica in situ. Posteriormente las bolsas fueron cerradas por medio de un sellador eléctrico (Orskov et al., 1980) y cada una de las bolsas fueron almacenadas en una malla de plástico de acuerdo al tiempo de incubación correspondiente y finalmente las mallas fueron atadas por medio de una cuerda de cáñamo a mosquetones fijados a una cadena de hierro cromado de 1.0 m de longitud.
Incubación de las bolsas
Justo antes de realizar la incubación ruminal, se enjuagaron las bolsas en agua a 39ºC durante 2 minutos (Giraldo et al, 2007), posteriormente se incubaron las bolsas en el rumen. Al momento de la incubación, se determinó el pH ruminal mediante un potenciómetro portátil (WTW 315l®). Se introdujo una bolsa por cada tiempo -0, 12, 24-, por cada tamaño de poro, por cada fuente mineral evaluada y en cada animal, de tal forma que se introdujeron un total de 24 bolsas por animal.
Las sales contenidas en las bolsas fueron introducidas al rumen al mismo tiempo en las tres vacas comenzando con el tiempo más largo -24 horas- prosiguiendo con las bolsas de los demás tiempos -12 y 0 h-, de tal manera que al final de la incubación, todas las bolsas fueron extraídas al mismo tiempo y se lavaron simultáneamente con agua de grifo hasta que el agua salió limpia (Nocek, 1988).
Determinación de la desaparición de la materia seca (DMS)
Con el objetivo de determinar la liberación o desaparición de la MS, las bolsas fueron secadas en una estufa de aire forzado a 65ºC durante 48 horas, luego de lo cual fueron pesadas. A dicho peso se le restó el peso de la bolsa vacía para calcular la cantidad de muestra desaparecida en cada tiempo de incubación. De esta forma la DMS parcial fue calculada como se muestra en la ecuación 1:
Análisis estadísticos
Se estableció el promedio y la desviación estándar del tamaño de partícula de cada fuente de Mg. Los datos de DMS fueron analizados bajo un diseño en bloques completos al azar con un arreglo factorial 2x3X4, donde los factores analizados fueron respectivamente, el tamaño del poro, el tiempo de incubación y las fuentes minerales. Las 3 vacas representaron los bloques, de acuerdo al modelo mostrado en la ecuación 2:
Para el análisis de los datos se utilizó el PROC GLM del programa estadístico SAS (2002) donde la variable respuesta a analizar fue la desaparición de la materia seca (DMS) expresada en %. Los resultados obtenidos fueron expresados como la Media ± Error Estándar. Cuando se hallaron diferencias significativas, se aplicó la prueba de LSMEANS para el análisis de la diferencia entre los promedios. Adicionalmente se adelantó un análisis de correlación entre la solubilidad de cada fuente y la DMS a la 24 h mediante el PROC CORR del programa estadístico SAS (2002).
Resultados y discusión
El pH promedio para los animales experimentales, medido antes de la introducción de las bolsas fue de 6,99±0,021. Dicho pH es compatible al hallado para animales pastando praderas de forrajes tropicales fibrosos y sin suplementación alimenticia (García & Kalscheur 2006, Correa et al, 2011, Duque 2013). Dicho valor (Ver Tabla 1) es importante puesto que este se asocia con la solubilidad de las fuentes inorgánicas de minerales para rumiantes (Jesee et al., 1980).
El tamaño promedio de partícula de las cuatro fuentes de Mg fue superior a 700 µm (Tabla 2), lo que puede indicar que la desaparición de las muestras de las bolsas de nailon no fue causada por el tamaño de partícula de la fuente mineral si no, por su solubilidad en el medio ruminal, toda vez que los tamaños de poro evaluados fueron 25 y 50 µm.
Tabla 2.
Tamaño de partícula para las fuentes minerales evaluadas.
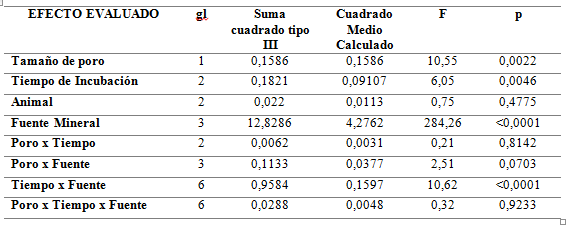
No se detectó efecto de la triple interacción entre la fuente, el tamaño del poro y el tiempo evaluado (p=0,92), como lo muestra la Tabla 3. Con respecto a la interacción de primer nivel, solamente se observó efecto de la interacción entre el tiempo de incubación en rumen y la fuente mineral (p<0,0001). Resultados similares son reportados por Correa (2007) en los que el Mg contenido en forraje kikuyo presentó una alta desaparición ruminal a las 0 horas (70%) y a las 24 horas alcanza una desaparición superior al 90%. Igualmente, Gojon et al. (1998) observaron que los principales macro minerales (Ca, P, Mg, K) presentaban a las 24 horas una desaparición ruminal in situ de aproximadamente 85%.
Tabla 3.
Efecto del tamaño del poro de la bolsa, fuente mineral y tiempo de incubación sobre la desaparición de materia seca DMS de las fuentes minerales evaluadas.
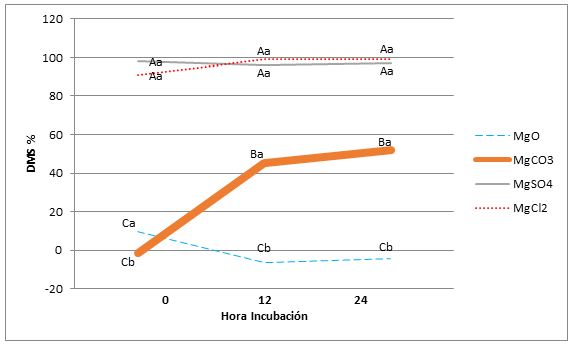
No se detectaron efectos significativos del tiempo de incubación sobre el nivel de DMS para MgCl2 ni para MgSO4 (p=0,74) (Gráfica 1). Estas fuentes fueron estadísticamente superiores en todas las mediciones a las otras dos fuentes evaluadas (p<0,0001). La descripción técnica del cloruro y el sulfato de Mg demuestra que estas dos fuentes presentaban los mayores valores de solubilidad (54,2 y 35,5 g/100mL, respectivamente) (Royal Society of Chemistry). La DMS del MgCO3 y del MgO fue baja en el tiempo 0, pero luego se incrementó la del MgCO3 mientras que la del MgO presentó valores negativos, indicando el atrapamiento de contenido ruminal.
Figura 1. Efecto de la fuente mineral y el tiempo de incubación sobre la DMS (%)
Literales diferentes en mayúsculas indican diferencias en el tiempo
Literales diferentes en minúsculas indican diferencias dentro de la fuente
Un análisis de correlación entre la solubilidad en medio acuoso y la DMS a las 24 h (r=0,93 y r=0,90 para las bolsas de 50 y 25 µm, respectivamente), mostró que existe una asociación alta entre estos dos parámetros y que, por lo tanto, a mayor solubilidad de la fuente, mayor es la desaparición de la muestra de las bolsas de nailon. Se ha descrito que tanto el tamaño de partícula (Quin et al., 1938) como la solubilidad (Ortega & Carranco, 1993) como factores que afectan la desaparición ruminal de la materia prima incubada en la bolsa de nylon. Es posible que bajo las condiciones del presente estudio el efecto ejercido por el tamaño de partícula sobre la desaparición ruminal sea dependiente de una característica inherente a la materia prima evaluada como la solubilidad.
Efecto del tamaño del poro
Los tamaños de poro utilizados en este estudio se encuentran dentro del rango recomendado por un gran número de autores (Van Hellen & Ellis, 1977; Uden & Van Soest 1984; Lindberg, 1984; Nocek, 1985, 1988; Vanhatalo, 1995; Hvelplund & Weisbjerg 2000). No obstante que el tamaño de las partículas de las fuentes evaluadas (Tabla 2) superó varias veces el tamaño de poro más grande evaluado en las bolsas (50 µm), dicho factor afectó la DMS de las fuentes, siendo estadísticamente menor con las bolsas de 25 µm (51,7%) (p<0,002) que con las de 50 µm (61,1%). En este sentido cabe anotar que una proporción de dichas partículas (ver anexos) tenían un tamaño inferior y suficiente para escapar por los poros y además, con la motilidad y la humedad ruminal, muchas partículas redujeron su tamaño, lo cual facilitó su salida. Otros autores también han reportado una reducción en la digestibilidad de las muestras incubadas cuando el tamaño del poro de la bolsa es menor. Lindberg & Knutsson (1981) obtuvieron disminuciones significativas en la DMS y desaparición de N al trabajar con tamaños de poro de 80 a 120 µm. En este mismo trabajo, los autores observaron que al utilizar poros más pequeños (10 - 36 µm) y aumentar el tiempo de remojo previo a la incubación (6 horas con agitación) se disminuía la DMS.
Giraldo et al. (2007) por el contrario, observaron una mayor digestibilidad in vitro de la materia seca en kikuyo determinada por la técnica del Daisy II® con tamaño de poro de 20 µm que con la técnica in situ con bolsas de nailon cuyo tamaño de poro era de 50 µm. Este último resultado, sin embargo, pudo deberse a la diferencia en la relación tamaño de muestra: área efectiva de la bolsa, que habría favorecido las muestras evaluadas por la técnica Daisy II®, como lo indica los trabajos de Goering & Van Soest, 1970. Asi mismo, Van Hellen & Ellis (1977) han relacionado la DMS con la relación entre el tamaño de la muestra ingresada en cada bolsa y el área de cada cara de la bolsa incubada.
Efecto del tiempo y la fuente sobre la DMS (%)
Hubo una disminución significativa de MgO en la DMS luego de la hora 0 de incubación (p<0,03), presentando incluso valores negativos a las 12 y 24 horas, que no fueron diferentes entre sí (p=0,79). En este sentido, Jesse et al. (1981) trabajaron con tamaños de partícula del MgO similar al del presente estudio, obteniendo un bajo nivel de DMS (<50%), asi mismo, Morgan (2011) experimentando con vacas lecheras fistuladas que presentaron un pH ruminal similar al del presente estudio (6,5) observaron una DMS baja incubando MgO durante 96 horas. Por ello al tener las condiciones experimentales bajo las cuales se trabajaron en el presente experimento se podría esperar una baja DMS ruminal.
Los valores bajos de DMS se correspondieron con los menores valores de solubilidad reportados para esta fuente (0,0086g/100mL) (De Baat, 1926). Los valores negativos hallados, podrían responder al ingreso de materiales propios del rumen a través de los poros, que aunado a la baja solubilidad del MgO, generaron valores negativos de DMS. Nocek (1988) indica que al trabajar con materiales con tamaño de partícula grande (>90µm) la DMS disminuía significativamente. Igualmente, Ceballos et al. (2008) indican como opción para evitar datos negativos de DMS incubar una bolsa vacía por cada tratamiento, no obstante, en el presente estudio no se realizó este procedimiento por no estar recomendado por la mayoría de la literatura relacionada con la técnica in situ.
El MgO utilizado en el presente estudio tenía un tamaño de partícula de 963,9 ± 317µm y las bolsas utilizadas tenían tamaños de poro de 25 y 50 µm de tal manera que es posible que debido a este impedimento físico la mayoría de las partículas de MgO no hayan abandonado las bolsas de nailon. El tamaño de partícula del MgO utilizado en el presente estudio (MgO técnico) fue superior al reportado previamente por Schaefer et al. (1982) y por Xin et al. (1988) quienes trabajaron con MgO comercial con tamaño de partícula de 0.09 y 3.7μm, respectivamente. Es posible que la diferencia entre este estudio y los citados previamente es que en dichos estudios el MgO hizo parte de una mezcla de alimentos que fue incubada en el rumen.
En reportes previos, Xin et al. (1988) midieron la solubilidad de MgO al introducir este compuesto con diferentes tamaños de partícula en soluciones acidas de HCl (pH entre 1,3 y 5,6) revelando que a menor tamaño de partícula (237µ) y a menor pH (1,3) la solubilidad de esta sal fue mayor. Numerosos autores han reportado para ganado Holstein en pastoreo sin suplementación valores de pH ruminal de entre 6,0 y 6,79 (Holden et al., 1994; Reis & Combs 2000, Bargo et al., 2002, García & Kalscheur 2006, Correa et al., 2012, Duque 2013), lo que coincide con los hallados en este trabajo.
La utilización de este tipo de materiales (específicamente MgO) se restringe a la adición del producto comercial a dietas total o parcialmente mezcladas como tal, sin ningún proceso de molido o de filtrado (Emery et al., 1965; Askew et al., 1971; Schaefer et al., 1982). En el presente experimento el MgO no fue molido o filtrado pudiendo presentar partículas que por su tamaño, no lograron solubilizarse en el rumen. Aún a las 24 horas post-incubación se registraron DMS negativas, por lo que se puede sospechar inicialmente, que el óxido ligo partículas del medio ruminal. A este respecto, las reglas de solubilidad coinciden en afirmar que los óxidos de un ion metálico no son solubles en agua y tienden a la formación de hidróxidos (Morgan, 2011).
Lough et al. (1990), por su parte, encontraron que la solubilidad del MgO es alta a las 24 horas en liquido ruminal cuyo pH inicial osciló entre 6,95 y 6,99. Dicha solubilidad fue de 91,8% sugiriendo que aún a pH altos, esta fuente puede ser muy soluble. Es necesario tener en cuenta, sin embargo, que la solubilidad calculada por estos autores, fue la proporción de Mg hallado en la fase liquida frente a la fase sólida, luego de centrifugar el líquido ruminal. Lamentablemente en este estudio no se reportó el tamaño de partícula con la que se trabajó por lo que no se puede hacer ninguna inferencia al respecto. Los resultados de estos autores sin embargo contradicen la baja solubilidad reportada para el MgO (Reference Tables for Physical Setting - Chemistry 2011.) sugiriendo que se pueda tratar más de un error metodológico, que de una solubilidad real.
Otros autores, han reportado valores de digestibilidad negativos para Mg (Emanuelle et al., 1991) y para el Ca (Emanuelle et al., 1991, Correa et al., 2008), asociados a la quelación de estos iones divalentes con la materia orgánica del rumen, particularmente con la fibra. Es probable, entonces, que parte del valor negativo de la DMS observada para esta fuente, fuera debido a la quelación de componentes orgánicos por parte del MgO.
El MgO ha sido utilizado en las dietas de vacas lecheras para prevenir problemas de acidosis ruminal (Erdman et al., 1980, 1982; Schaefer et al., 1981) gracias a su capacidad de consumir iones H+ y por tanto, de formación de enlaces con otros compuestos. No obstante, de acuerdo a los resultados observados en el presente estudio, se puede esperar que la solubilización de este material a nivel ruminal sea menor que las demás fuentes evaluadas ya sea por su baja solubilidad en medios acuosos, por el pH presente en el rumen o por el impedimento físico del tamaño del poro utilizado.
Por el contrario, en el MgCO3 se observó un aumento significativo (p<0,001) y constante en la DMS a partir del tiempo 0 de incubación hasta las 24 horas. La DMS a las 24 h hallada en esta fuente puede estar relacionada con su naturaleza química, ya que al ser una sal anhidra presenta una solubilidad muy superior que la del MgO.
De acuerdo a los valores reportados en la Tabla 2, la solubilidad de este carbonato sería 1200 veces superior a la del óxido. Sin embargo, todas las formas de MgCO3 son poco solubles en condiciones neutras y básicas e incrementan su solubilidad en medios que tienden a la acidez (Patnaik & Pradyot 2003). En este sentido Scandolo et al. (2007) observaron una disminución constante del pH ruminal a lo largo del día en vacas lecheras. De esta forma, la DMS del MgCO3 registrada en el presente experimento siguió el comportamiento esperado, toda vez que aumentó a lo largo del día -las bolsas fueron incubadas a las 08:00- es decir la DMS aumentó a medida que disminuye el pH ruminal coincidiendo con las observaciones realizadas por dichos autores. Bajo las condiciones de pH del ambiente ruminal de las vacas experimentales trabajadas en este ensayo, era de esperarse una baja solubilidad, aunque mayor que la del MgO, tal y como se encontró.
Efecto del tamaño de partícula de las fuentes evaluadas
El tamaño de partícula del MgCO3 utilizado en el experimento fue de 740 μm, menor al del MgO; a pesar de esto, la consistencia del carbonato era tal que al entrar en contacto con una solución acuosa, producía una masa de disolución lenta lo que pudo disminuir la liberación inicial del material a través de los poros. No obstante, no se observaron valores negativos de DMS y por el contrario este parámetro aumentó significativamente hasta alcanzar aproximadamente un 50% de DMS a las 24 horas de incubación.
El MgSO4 y el MgCl2 presentaron los valores más altos de DMS sin que hubiese diferencias entre ellos durante los tres periodos de incubación ruminal. Esto puede ser debido a la mayor solubilidad de estas dos fuentes comparadas con la del MgO y la del MgCO3 (Tabla 2). La diferencia en la solubilidad entre estas dos fuentes (35,5 y 54,2 g/100 mL para el MgSO4 y el MgCl2, respectivamente), no influyó en la DMS en ninguno de los tiempos de incubación debido a que la cantidad necesaria para precipitarse (17750 y 27100 g en 50 L de líquido ruminal), es mucho mayor que la que se incubó en las bolsas experimentales (1,5 g en 50 L de líquido ruminal).
Dentro de las fuentes evaluadas, el MgSO4 presentó el mayor tamaño de partícula (992 μm ver anexos), no obstante, al ser un compuesto altamente higroscópico (Hutjens, 1991), incluso a temperatura ambiente, la absorción de agua por parte del material, disminuía notablemente el tamaño de las partículas que lo componían, facilitando su liberación a través de los poros.
El MgSO4 se ha utilizado rutinariamente para corregir deficiencias de magnesio en cultivos de algunas hortalizas y cereales (Peña et al., 2007), como tranquilizante en cerdos (Panella-Rivera et al., 2009), en ratones para evitar daños renales (Masuyama et al., 1995) y en vacas lecheras con problemas de hipomagnesemia (Wittwer et al., 1995). En el caso específico de vacas lecheras algunos autores (Capen & Rosol, 1989; Hoffsis et al., 1989; Xin et al., 1989; Contreras, et al., 1992) han afirmado que el suministro oral de MgSO4 funciona en el tratamiento de vacas con hipomagnesemia, aumentando los niveles séricos del mineral.
En el presente experimento se utilizó MgCl2 de grado técnico -con textura y granulometría diferentes al comercial-, que presentó el menor tamaño de partícula de todas las fuentes evaluadas (700 μm, Tabla 2). Esta característica, sumada al hecho de que este material fuera higroscópico, hizo que su liberación y sobre todo su solubilización fueran más rápidos que en las demás fuentes minerales evaluadas.
En un estudio de cinética de liberación de pasto Kikuyo (Correa, 2007) trabajando con una metodología similar, se observó que la liberación ruminal del Mg presente en el forraje era inicialmente rápida, 70 - 80% en las primeras dos horas, para aumentar hasta las 24 horas sin que mostrará un incremento adicional durante las siguientes 48 horas de incubación, un patrón de comportamiento similar al observado con el MgSO4 y el MgCl2, en el presente estudio aunque de menor magnitud. Sin embargo, la solubilidad del Mg contenido en sales puras es más extensa y rápida que la reportada en el Mg contenido en forrajes, teniendo en cuenta que la naturaleza inorgánica de las sales puras influye en la solubilidad. Cole (2000) por otro lado, observó luego de 30 minutos del consumo de forrajes tiernos y concentrados, un incremento significativo de la concentración ruminal de Mg.
Normalmente la concentración de Mg en el fluido ruminal puede cambiar rápidamente a lo largo del día, dependiendo del suministro dietario del mineral (Scandolo et al., 2007). Por ello, al contar con una liberación rápida y alta de Mg en el rumen como sucede con el MgCl2 y el MgSO4, se puede esperar que su función como estimulante en la actividad microbiana (Galbraith et al., 1971; Rusell & Dombrowski, 1980), bufferizante (Rauch et al., 2012; Erdman et al., 1982, 1988; Xin et al., 1989) y como cofactor en la actividad de algunas enzimas (Faixová & Faix, 2002; Faixová et al., 2006) sea rápida luego de su consumo y que no se mantenga en el tiempo.
Como se pudo establecer, las fuentes de Mg evaluadas en este trabajo presentaron patrones de DMS diferentes indicando que su papel en el suministro de este mineral es igualmente distinto y que, por lo tanto, una mezcla de estas fuentes podría tener un efecto más positivo sobre el crecimiento microbiano y el cubrimiento de las necesidades del animal hospedero que una sola fuente de lenta, media o alta disponibilidad. Este sería un enfoque similar al que se ha propuesto para carbohidratos y proteínas en la elaboración de suplementos para rumiantes (Rotger et al., 2006; Cole & Todd 2007; Hall & Huntington, 2007). Bajo este enfoque y haciendo parte de una mezcla en la que también estén presentes carbohidratos con diferente capacidad de fermentación, los cambios en el pH, ocasionados por la degradación de los carbohidratos y las proteínas podrían favorecer tanto la liberación del Mg de estas fuentes como su absorción ruminal.
Conclusiones
La desaparición de la materia seca fue afectada por el tamaño del poro y por la interacción entre el tiempo de incubación y la fuente de Mg evaluada (Ver Tabla 2). A pesar de que el tamaño de partícula de todas las fuentes inorgánicas fue mucho mayor al de los tamaños de poro evaluados, la DMS de las muestras de las bolsas de nailon no fue afectada por el tamaño de partícula si no, por el tamaño del poro de las bolsas y por la solubilidad de la fuente en el medio ruminal.
La DMS registrada en las bolsas de 25µm fue inferior a la observada en las de 50 µm siendo un efecto esperado y reportado previamente en la literatura. Para algunas fuentes minerales se observó alta variabilidad en el tamaño de partícula, de tal manera que una proporción de partículas con tamaño inferior pudieron escapar por los poros y otras redujeron su tamaño por efecto de la motilidad y la humedad ruminal. La relación entre la cantidad de muestra y el área efectiva de la bolsa fue exactamente la misma en las bolsas Brioni® y las ANKOM®.
El MgO presentó la menor DMS a las 24 horas, registrando incluso valores negativos que estarían asociados con el tamaño de partícula, el pH ruminal de los animales experimentales y el bajo nivel de solubilidad en medio acuoso de esta sal. De igual forma, las propiedades inherentes de los óxidos de metales divalentes y su relación con la posible formación de quelatos, pudo influenciar la DMS con valores negativos.
La consistencia del MgCO3, el nivel medio de solubilidad y el pH ruminal causaron que esta fuente mineral haya alcanzado una DMS final menor a 50%.
La DMS en todos los tiempos evaluados para el MgCl2 y el MgSO4 fue similar. No obstante, la DMS de estas dos fuentes fue superior a las demás sales de Mg durante los tres tiempos de incubación. Es posible que el lavado previo con agua de grifo y la naturaleza higroscópica de estas dos fuentes, hayan generado un valor de DMS superior en el primer tiempo de incubación.
La utilización de MgCl2 o de MgSO4 como fuentes inorgánicas en dietas para rumiantes garantiza una alta y rápida liberación de Mg debido a la mayor solubilidad de estas sales y por tanto, mayor disponibilidad para los microorganismos y la mucosa ruminal. Sin embargo, a nivel práctico es probable que la mezcla de fuentes que presenten un patrón acelerado de desaparición ruminal, que pueda ser más apropiada para cubrir las necesidades de las poblaciones microbianas y del animal hospedero.
Agradecimientos
Los autores agradecen al zootecnista Santiago Jaramillo de la Universidad Nacional sede Medellín, al estudiante de Maestría Cesar Orrego y a la Ingeniera Carol Lizzeth Martínez por su colaboración en el trabajo de campo, la toma de datos y el procesamiento de las muestras. Al laboratorio de Nutrición Animal de la Universidad Nacional de Colombia Sede Medellín por su colaboración con las instalaciones y los equipos necesarios para el desarrollo de este experimento. Al centro de producción de Paysandú, en especial a Neftalí Ortiz por el apoyo logístico y la colaboración con los animales experimentales.
Literatura Citada
ANEXOS
Muestras de partícula para la determinación del tamaño de partícula de las fuentes minerales utilizadas en el experimento, medidas sobre papel milimetrado con divisiones de 1mm (1000µm)
1 Tamaño de Partícula MgO

2 Tamaño de partícula MgCO3

3 Tamaño de partícula MgSO4

4 Tamaño de partícula MgCl2